摘要
钙粘蛋白/连环蛋白复合体在发育、分化、缺血和神经退行性疾病(如阿尔茨海默病(AD))中通过细胞-细胞相互作用调节多种信号通路。几种影响钙粘蛋白介导的信号通路的刺激可调节跨膜蛋白的处理,包括淀粉样前体蛋白(APP),下调wnt相关信号通路,或促进核转录程序。核磷酸肌醇信号通路调节应激介导的染色质修饰,促进细胞极性或恶性肿瘤的发展。有趣的是,血管内皮(VE)-钙粘蛋白/连环蛋白复合物可以调节缺氧或中风引起的炎症条件下的内皮屏障功能和血管稳定性。血脑屏障的通透性和营养物质的获取是脑功能正常的前提。中风、缺氧缺血脑病的发生破坏了由ve -钙粘蛋白/血管内皮生长因子(VEGF)受体信号调节的内稳态,这一过程在成人和新生儿的大脑中是不同的。n -钙粘蛋白和VEGF调节血管生成或血管反应以及神经祖细胞向脱髓鞘病变的募集或迁移。缺氧诱导因子(HIF)信号轴激活了心室下区(SVZ)或皮层下白质道内皮细胞和少突胶质前细胞(OPCs)中netrin-1和wnt介导的连环蛋白信号等关键分子,HIF信号的时空失调可能是心室周围白质软化的主要原因。本文就钙粘蛋白/连环蛋白介导的机制和相关的病理障碍进行了综述,重点介绍了钙粘蛋白/连环蛋白复杂分子框架的组织以及钙粘蛋白/连环蛋白复合物介导的几种发育功能。
关键字
钙粘蛋白,缺氧,p120连环蛋白,室周白质软化,早衰素
简介
钙粘蛋白/连环蛋白细胞黏附复合物是突触发生、可塑性、内皮存活和血管形态发生所必需的[1,2]。这些细胞粘附机制受多种连环蛋白调控,包括a-、b-和d-,或在其中心结构域具有armadillo (Arm)重复的p120连环蛋白。a-Catenin与多种肌动蛋白结合蛋白如a-肌动蛋白相互作用,调节肌动蛋白细胞骨架的动态。它的相互作用伙伴,b-catenin,通过与经典的E-或N-cadherins[3]相互作用,构成钙粘蛋白介导的粘附连接。b-Catenin还通过与pdz结构域蛋白结合调节突触内稳态,并调节神经元[4]中的神经元活性或树突形态发生。p120连环蛋白通过近膜(JMD)序列与经典钙粘蛋白结合,并通过早老素-1 (PS1)调节其处理[5,6]。在速度心面肌综合征(ARVCF)亚家族中缺失的p120连环蛋白和犰狳重复蛋白都可以作为两种主要的剪接异构体存在,其区别在于n端存在(异构体1)或不存在(异构体3)盘绕圈区,并调节质膜[7]中的经典钙粘蛋白运输和稳定性。有趣的是,δ相互作用蛋白A (DIPA)被鉴定为1亚型特异性p120结合蛋白。DIPA的敲低和过表达引起的表型类似于n-钙粘蛋白突变体中看到的表型,即脑积水和异位,这与大脑发育[8]期间钙粘蛋白介导的异常信号有关。此外,p120连环蛋白参与了ps1介导的cadherin加工过程,该过程可与g-secretase复合物组织的APP切割进行上下文依赖性的竞争[6]。 In addition, nuclear phosphatidylinositol-4,5-bisphosphate PI(4,5)P2-mediated signaling is necessary for the regulation of transcription and promotes E-cadherin biogenesis or suppresses inhibitor of growth protein 2 (ING2)-mediated chromatin remodeling in response to several stimuli [9].
在缺氧条件下,HIF-1通过抑制其亚基HIF-1a的蛋白酶体降解来稳定,而已知PS1和PS2都调节HIF-1a的周转[10]。PS1和PS2促进缺氧依赖性HIF-1a的表达,这种表达依赖于APP或g-secretase生成的其胞内结构域(AICD)。另一方面,HIF-1信号在脑中动脉闭塞(MCAO)后脑梗死或新生儿卒中中检测到血管内皮生长因子(VEGF)上调,VE-cadherin是VEGF介导的血管生成所必需的[1,11]。VEGF通过刺激b-阻滞素依赖的ve -钙粘蛋白[12]的内吞作用来控制内皮细胞的通透性。有趣的是,缺血性中风后血脑屏障功能的破坏在新生儿和成人中是不同的。事实上,与中风后的成人大脑相比,未成熟大脑中紧密连接蛋白的表达在很大程度上保留了下来,而一些增加血管通透性的基因,如VEGF受体2,只在成人大脑中过表达[13,14]。
来自SVZ的胶质纤维酸性蛋白(GFAP)阳性星形细胞可产生神经元、ng2阳性少突胶质细胞祖细胞(OPCs)和成熟的髓鞘化少突胶质细胞[15]。通过与白质血管生成耦合,opc固有的HIF1/2a是出生后髓鞘形成所必需的,如条件性HIF1/2a敲除小鼠前脑白质束丢失所示,其表型使人联想到被分类为早产儿[16]的脑室周围白质软化(PVL)。根据皮层下白质脱髓鞘模型,SVZ中神经元祖细胞(NPCs)的扩张和NPCs中分解素和金属蛋白酶(ADAM)蛋白酶激活n -钙粘蛋白信号都是增强NPCs向脱髓鞘病变[17]迁移的必要条件。缺氧缺血导致白质损伤导致OPC扩大,SVZ祖细胞在恢复期死亡似乎是缺氧缺血性脑病(HIE)或新生儿缺氧的共同特征[17-20]。本文就发育障碍和新生儿缺氧过程中钙粘蛋白依赖性细胞/细胞通信的信号通路进行了综述。
钙粘蛋白/连环蛋白复合物的分子组织
钙粘蛋白家族蛋白根据其结构和功能分为几个亚组。经典的钙粘蛋白是I型跨膜蛋白,其胞外结构域包含5个EC子结构域(EC1-EC5),介导同质性相互作用的稳定通过Ca2 +绑定[2]。它们的细胞质结构域包含与p120连环蛋白结合所需的近膜(JMD)序列,并在c端与b-连环蛋白相互作用。b-Catenin通过与a-catenin相互作用调节肌动蛋白的细胞骨架,EPLIN(肿瘤中丢失的上皮蛋白)和Afadin介导F-actin与钙粘蛋白/catenin复合物之间的连接[21,22]。p120连环蛋白家族由p120连环蛋白、ARVCF、d-连环蛋白/NPRAP、血小板蛋白和p0071组成。p120 catenin和ARVCF都普遍存在于两种具有或不具有n端调节卷曲结构域(异构体1和3)的剪接变体中。ARVCF、d-catenin和p0071在突触或棘中具有部分重叠功能,并在c端结构域[7]具有PDZ结构域。d-Catenin敲低对脊柱和突触密度的影响与钙粘蛋白结合无关,这表明PDZ区域的生物学重要性。p120连环蛋白在钙粘蛋白和Rho家族蛋白之间具有协调作用,调节脊柱和突触的形态发生[23]。p120连环蛋白在钙粘蛋白加工过程中也起着关键作用,并与APP加工过程竞争,APP加工过程是由含有早老素-1 (PS1)[6]的有限g-secretase复合物介导的。
p120异构体1 (p120-1)包含一个由拼接产生的n端螺旋结构域,主要表达于间充质系或神经元中(图1)。相比之下,p120异构体3 (p120-3)主要表达于上皮细胞中。上皮-间质转化(EMT)与p120-3或4 - p120-1交替转换和经典的E-向n -钙粘蛋白转化[24]协调。DIPA家族蛋白特异性结合p120-1,但不结合p120-3,并与粘附连接共定位。有趣的是,DIPA的低表达或过表达导致神经管闭合和神经上皮组织紊乱的缺陷,类似于斑马鱼[8]中n -钙粘蛋白突变的模型。含有蛋白85C (Ccdc85c)的线圈结构域,一种DIPA家族蛋白,已知在出血性脑积水中被破坏(hhy)突变小鼠模型[25]。因为Ccdc85c被招募到粘附结通过p120-1,提示Ccdc85c/p120连接缺陷导致皮质下异位和脑积水。
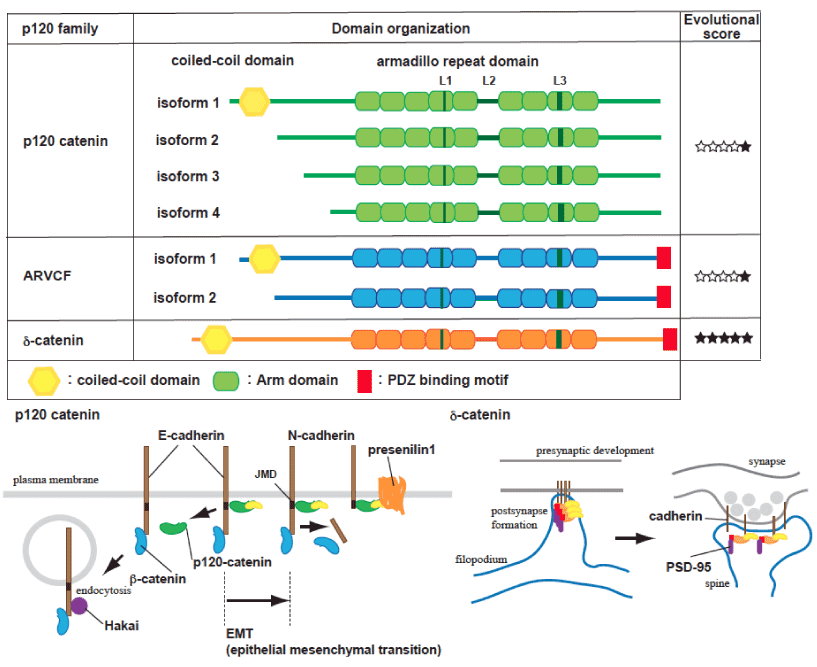
图1。p120连环蛋白家族蛋白示意图。
p120连环蛋白基因包含4个不同的翻译起始位点(isoform 1-4),在n端和c端通过选择性剪接产生多个蛋白变体。大多数细胞表达多种形式的p120连环蛋白,都具有9个中央串联重复的Armadillo结构域,L结构域(L1-L3)和中间和c末端的外显子交替拼接(A-C)。p120连环蛋白亚型1在n端包含一个螺旋结构域和多个磷酸化区域。p120连环蛋白异构体3和4缺少螺旋结构域和大部分n端调控结构域,主要在上皮细胞中表达。n端调控结构域具有磷酸化位点,是受体酪氨酸激酶诱导的响应表皮生长因子和肝细胞生长因子的细胞运动所必需的。ARVCF有两种主要的异构体,如p120 catenin, ARVCF和d-catenin在c端尾部包含一个PDZ结构域,该结构域介导与PSD-95[47]的相互作用。填充的星形符号表示在系统发育史中存在相应的p120连环蛋白家族的同源物,被标记为棘突动物、刺胞动物、原气孔动物、脊索动物和脊椎动物(7)。钙粘蛋白复合体在质膜的动态平衡下调节,依赖于与p120连环蛋白家族蛋白的关联。通过APP处理平衡中的限制性蛋白水解[6],钙粘蛋白的周转也需要PS1。N-向e -钙粘蛋白的转化发生在上皮-间质转化(EMT)过程中,p120 catenin未结合的钙粘蛋白靶向内化并受hakai介导的降解[28,30,32]。A -和b-连环蛋白在聚集钙粘蛋白和调节细胞骨架动力学方面具有重要作用,但它们对表面钙粘蛋白水平的影响被认为是间接的。 d-catenin, which can be localized in dendrites or synapses, is involved in the regulation of PSD95 and N-cadherin levels, Interaction with the PDZ domain binding protein, but not cadherin binding, is necessary for activity-dependent d-catenin-mediated signal [48].
病理状态下钙粘蛋白/连环蛋白的功能
室管膜前体在胎儿室壁表面产生放射状胶质细胞和纤毛室管膜细胞;然而,hhy突变脑表现为室管膜发育不全伴炎性变性[25]。n-钙粘蛋白黏附连接的破坏导致室管膜凋亡和脑室壁[26]的剥蚀。在大脑发育过程中,p120连环蛋白和n -钙粘蛋白在增生性SVZ的神经母细胞中共定位。E-cadherin/b-catenin分子机制功能障碍导致的神经母细胞迁移缺陷可以在hyh携带错义突变的突变体(脑积水伴跳跃步态)纳帕该蛋白编码可溶性n -乙基马来酰亚胺敏感因子附着蛋白α (aSnap),并影响脑室神经上皮[27]的组织。
p120连环蛋白通过阻止胞吞作用参与了质膜上经典钙粘蛋白的稳定,在癌症进展到[28]转移期期间,p120连环蛋白的大量丢失导致e -钙粘蛋白的丢失。相比之下,Numb通过直接与NVYYY基序相互作用结合p120/E-cadherin复合体,促进E-cadherin的内吞[29]。E-cadherin中的基序被Src激酶磷酸化,并募集c- cbl相关的E3泛素连接酶Hakai,导致内吞和降解[30-32]。另一方面,e -钙粘蛋白的生物生成受多种与内在或外源性刺激相关的信号机制调控[9,33]。维生素D受体配体,1a,25-二羟基维生素D3 (1a,25(OH))2D3.),激活RhoA和p38丝裂原激活蛋白激酶(p38 MAPK),伴Ca2 +E-cadherin表达[33],促进分化。相比之下,Ca2 +-诱导机制,核PI(4,5)P2在使用维生素D受体配体1a,25-二羟基维生素D3 (1a,25(OH)处理后,发现由磷脂酰肌醇-5-磷酸4激酶II型β (PIPKIIb)催化生成的产物对结肠癌模型中E-cadherin的上调至关重要。2D3.磷酸肌醇苷5 ' -磷酸(PI(5)P)与ING2相互作用,ING2是一种肿瘤抑制蛋白,含有植物同源结构域指基motif,在染色质调节蛋白中非常常见,ING2促进应激诱导的细胞死亡通过p53激活途径[34]。ING2通过与PI(5)P直接结合,组成染色质调节复合体的一部分,以应对遗传毒性应激。这种ING2/PI(5)P相互作用是稳定启动子占用的先决条件,允许调节细胞周期进展[35]。这些结果表明核PI(4,5)P2PI(5)P可能通过感应外界信号来控制细胞的生长和粘附性能。

图2。PI(5) p4 -激酶II对SW480细胞的上皮样极化b-介导的核PI(4,5)P2信号。
SW480细胞是人结直肠癌模型,用于研究肿瘤进展的表型变化和上皮样极化模型[49]。缺乏维生素D受体表达的SW480R细胞(左)呈圆形形态,生长快速,不依赖于锚点,失去接触抑制和粘附。相比之下,SW480-ADH细胞(中)保留了配体(1a,25(OH)2D3)激活的VDR激活特性,并获得E-cadherin表达的上皮形态。磷脂酰肌醇5-磷酸4激酶II型β (PI(5) p4激酶IIb)在VDR激活时介导核PI(4,5)P2的生成,是SW480-ADH细胞[9]中E-cadherin转录上调所必需的。
血管生成素样4 (Angtpl4)由缺氧诱导,并表现出独立于VEGF[36]的促血管生成反应。Angtpl4诱导缺血性脑卒中模型梗死区域明显减少,可能是由于VE-cadherin完整性的保留和PECAM稳定性的增加保护了血管完整性。miR-27a被鉴定为特异性靶向VE-cadherin,已知miR-27a的下调是血管生成所必需的,与miR-27抑制剂对VEGF反应中血管通透性的降低相一致,这表明通过转录调控[37]对VE-cadherin/VEGF信号通路进行缺氧依赖性调控的重要性。
钙粘蛋白介导的缺氧/缺血反应信号
HIF-1和HIF-2是由O组成的转录异质二聚体2-调控的HIF-1a或HIF-2a亚基和组成稳定的HIF-1b[38]。在O2,两个HIF-a亚基都受到脯氨酰-4-羟化酶结构域(PHD)蛋白的羟化作用,这导致它们的蛋白酶体降解。然而,在缺氧条件下,羟化过程被抑制,hif形成稳定的复合体,激活缺氧反应基因的转录,如VEGF或BNIP3。缺氧依赖性HIF-1a的表达由PS1和PS2以及严格调控的PHD2表达共同促进。HIF-1a的诱导依赖于g-secreatse产生的APP或AICD,而不依赖于Notch细胞内结构域(NICD)介导的转激活[10]。有趣的是,少突胶质前体细胞(OPCs)的发育需要大量的营养需求,将大量髓鞘片段包裹到轴突上,HIF1/2a的激活严格调节血管生长中wnt7a/b介导的b-catenin信号通路,形成白质束[16]。
VEGF是血管生成的主要调节因子,位于成人SVZ的血管作为血管龛[39]调节神经发生和神经母细胞迁移。据报道,在大脑中动脉闭塞(MCAO)[40]模型中,SVZ神经母细胞对缺血应激表现出增殖反应,并伴随迁移特性的丧失。脑卒中后室管膜细胞诱导胶质纤维酸性蛋白(GFAP)和血管网络。大脑中动脉[11]永久闭塞,侵袭性血管生长,缺血边界区VEGF和VEGF受体表达上调。延长脑室内VEGF供应可促进血管生成和脑内皮细胞周细胞覆盖,增加脑血流通过局灶性脑缺血[41]时脑微血管n -钙粘蛋白的上调。
通过在转基因小鼠[42]的大脑中表达诱导分泌的VEGF诱骗受体表明,特定的VEGF依赖性血管生成期对心室周围血管和功能性血脑屏障的发育和维持非常重要。在不改变局部血流[43]的情况下,抗vegf治疗缺血急性期血管功能障碍可有效阻断水肿形成和组织损伤。有趣的是,在新生儿缺氧脑病模型中,中风后血管生成明显延迟,中风[44]后缺血周围区域内皮细胞增殖没有受到刺激。此外,中风后新生儿血脑屏障增加,中性粒细胞转运受到抑制,这种不同的反应与基底膜组分上保留的分子成分和新生儿[13]损伤区域的粘连分子如层粘连蛋白和闭塞带蛋白1有关。
在脱髓鞘模型中,饲粮铜硫酮喂养的小鼠皮质下白质出现慢性脱髓鞘,而溶血卵磷脂治疗诱导胼胝体[17]发生急性局灶性脱髓鞘。在这两种动物模型中,在静息的gfap阳性神经干细胞和egfr阳性扩增npc中,均检测到ADAM10和N-cadherin及其结合蛋白b-catenin和p120 catenin的上调。从SVZ小生境迁移的egfr阳性npc具有较高的n -钙粘蛋白处理活性,并以adam10依赖的方式被招募到病变中。gfap阳性的初级祖细胞(B型)穿透室管膜层,也产生ng2阳性的OPCs和髓鞘化少突胶质细胞。脱髓鞘病变中,NG2迁移+EGFR表达明显高于皮层非迁移NG2+细胞[15 20]。祖细胞募集的病理反应似乎需要与血管反应的物理联系,netrin-1和VEGF已被确定为这一过程中的关键分子[19]。一个相似的潜在机制可能调节脑室周围白质软化[45]的病理过程。胚胎期VEGF阻断可影响外侧神经节隆起损伤后的神经元迁移,但不影响兴奋性锥体神经元的层压。这表明由于心室周围血管的丧失而引起的原发性心室周围病变可诱导类似PVL表型的继发性神经病理[42,46]。
结论
钙粘蛋白/连环蛋白系统由许多相互作用的蛋白质调节。该系统对环境刺激的反应控制着突触功能、血管发育和内稳态,也通过细胞/细胞通信建立细胞极性。钙粘蛋白的表达和稳定性由核脂质信号介导的PI(4,5)P调节2和mirna介导的信号通路。p120连环蛋白调节几个过程,包括通过钙粘蛋白结合和蛋白水解ps1介导的APP处理,以及缺血性损伤中神经祖细胞的迁移。HIFs介导几种缺氧反应,包括大脑发育过程中vegf介导的血管生成。新生儿缺氧的修复过程或脑室周围发病机制需要几个N-或ve -钙粘蛋白和基板组分介导的信号传导过程。分子途径似乎不同于成人的发病机制,阐明由钙粘蛋白控制的潜在机制可以创造治疗工具,以避免新生儿的神经元死亡和骨髓缺失。
确认
我感谢纽约西奈山医学院的Robakis NK博士允许我提及我们合作工作的总结。
参考
- Carmeliet P, Lampugnani MG, Moons L, Breviario F, Compernolle V等(1999)小鼠ve -钙粘蛋白基因的靶向缺失或胞质截断会损害vegf介导的内皮细胞存活和血管生成。细胞98: 147 - 157。[Crossref]
- 欧阳明,陆松,Kim T, Chen CE, Seong J,等(2013)N-cadherin通过独特的β- catenin依赖的P120ctn和β- catenin信号通路调节空间极化信号。Nat Commun4: 1589。
- Fannon AM, Colman DR(1996)基于钙粘蛋白粘附特异性差异的中枢突触连接复合体形成模型。神经元17: 423 - 434。[Crossref]
- Yu X, Malenka RC(2003) -连环蛋白对树突形态发生至关重要。Nat >6: 1169 - 1177。[Crossref]
- Piedra J, Miravet S, Castaño J, Pálmer HG, Heisterkamp N,等(2003)p120连环蛋白相关的Fer和Fyn酪氨酸激酶调节-连环蛋白tyr142磷酸化和-连环蛋白- α -连环蛋白相互作用。摩尔细胞生物23日:2287 - 2297。[Crossref]
- Kouchi Z, Barthet G, Serban G, Georgakopoulos A, Shioi J等(2009)p120 catenin招募钙粘蛋白到γ -分泌酶并抑制Abeta肽的产生。J临床生物化学284: 1954 - 1961。[Crossref]
- Carnahan RH, Rokas A, Gaucher EA, Reynolds AB (2010) p120-连环蛋白亚家族的分子进化及其功能联系。《公共科学图书馆•综合》5: e15747。[Crossref]
- Markham NO, Doll CA, Dohn MR, Miller RK, Yu H,等(2014)dipa家族螺旋-线圈结合p120-连环蛋白家族保守的异构体特异性头部结构域:在脑积水和异位中的潜在作用。细胞杂志25日:2592 - 2603。[Crossref]
- 口内Z,藤原Y,山口2021版权所有。SW480细胞中维生素D受体依赖的E-cadherin表达需要II型激酶。生物化学生物物理学研究408: 523 - 529。[Crossref]
- Kaufmann MR, Barth S, Konietzko U, Wu B, Egger S,等。(2013)早衰素/ γ -分泌酶功能缺失突变导致缺氧诱导因子失调。J >33: 1915 - 1926。[Crossref]
- Marti HJ, Bernaudin M, Bellail A, Schoch H, Euler M.等(2000)脑缺血后缺氧诱导的血管内皮生长因子表达先于新生血管。是中草药156: 965 - 976。[Crossref]
- Gavard J, Gutkind JS (2006) VEGF通过促进-抑制素依赖性的ve -钙粘蛋白内吞来控制内皮细胞通透性。Nat细胞生物8: 1223 - 1234。[Crossref]
- Fernández-López D, Faustino J, Daneman R, Zhou L, Lee SY,等(2012)成年急性脑卒中大鼠血脑屏障通透性增加,但新生儿脑卒中大鼠血脑屏障通透性不增加。J >32: 9588 - 9600。[Crossref]
- Bouleti C, Mathivet T, Coqueran B, Serfaty JM, Lesage M,等。(2013)血管生成素样4对缺血性卒中脑血管及功能损伤的保护作用。心欧元J34: 3657 - 3668。[Crossref]
- Menn B, Garcia-Verdugo JM, Yaschine C, Gonzalez-Perez O, Rowitch D,等(2006)成人大脑室下区少突胶质细胞的起源。J >26日:7907 - 7918。[Crossref]
- Yuen TJ, Silbereis JC, Griveau A, Chang SM, Daneman R,等(2014)少突胶质细胞编码的HIF功能与出生后髓鞘形成和白质血管生成相关。细胞158: 383 - 396。[Crossref]
- Klingener M, Chavali M, Singh J, McMillan N, Coomes A, et al. (2014) N-cadherin促进神经祖细胞从SVZ神经干细胞龛募集和迁移到脱髓鞘病变。J >34: 9590 - 9606。[Crossref]
- 罗丽娜,万燕,王晓燕,等。(2008)慢性围产期白质损伤对少突胶质细胞谱系成熟的抑制作用。安神经63: 520 - 530。[Crossref]
- Cayre M, Courtès S, Martineau F, Giordano M, Arnaud K, et al. (2013) Netrin 1有助于脱髓鞘后心室下区血管重塑和促进祖细胞迁移。发展140: 3107 - 3117。[Crossref]
- Aguirre A, Rizvi TA, Ratner N, Gallo V(2005)过度表达表皮生长因子受体赋予非迁移性出生后神经祖细胞迁移特性。J >25日:11092 - 1110。[Crossref]
- Abe K, Takeichi M (2008) EPLIN调节钙粘蛋白连环蛋白复合物与f -肌动蛋白的连接,并稳定环向肌动蛋白带。美国国家科学研究院105: 13 - 19。[Crossref]
- Sawyer JK, Harris NJ, Slep KC, Gaul U, Peifer M(2009)果蝇的afadin同源物Canoe在顶端收缩过程中调节肌动蛋白细胞骨架与粘附连接的连接。J细胞186: 57 - 73。[Crossref]
- Elia LP, Yamamoto M, Zang K, Reichardt LF (2006) p120连环蛋白通过rho家族gtpase和钙粘蛋白调节树突棘和突触的发育。神经元51: 43-56。[Crossref]
- Yanagisawa M, Huveldt D, Kreinest P, Lohse CM, Cheville JC,等(2008)p120连环蛋白亚型开关影响Rho活性,诱导肿瘤细胞侵袭,并预测转移性疾病。J临床生物化学283: 18344 - 18354。[Crossref]
- Mori N, Kuwamura M, Tanaka N, Hirano R, Nabe M,等(2012)在出血性脑积水(hhy)小鼠中编码径向胶质细胞顶端连接蛋白的Ccdc85c被破坏。是中草药180: 314 - 327。[Crossref]
- Oliver C, González CA, Alvial G, Flores CA, Rodríguez EM, et . (2013) CDH2/ n- cadherin基粘附连接的破坏导致室管膜细胞凋亡和脑室壁剥离。神经病理学杂志72: 846 - 860。[Crossref]
- Chae TH, Kim S, Marz KE, Hanson PI, Walsh CA (2004) hyh突变揭示了α Snap在顶端蛋白定位和控制神经细胞命运中的作用。Nat麝猫36: 264 - 270。[Crossref]
- Thoreson MA, Reynolds AB(2002)人类癌症中连环蛋白p120表达的改变:对肿瘤进展的影响。分化70: 583 - 589。[Crossref]
- 王铮,Sandiford S,吴超,李ss (2009) Numb对酪氨酸激酶信号的响应调节细胞粘附和极性。EMBO J28日:2360 - 2373。[Crossref]
- Fujita Y, Krause G, Scheffner M, Zechner D, Leddy HE, et al. (2002) Hakai,一种c- cbllike蛋白,泛素化并诱导e -钙粘蛋白复合体的内吞作用。Nat细胞生物4: 222 - 231。[Crossref]
- Palacios F, Tushir JS, Fujita Y, D’souza - schorey C(2005)溶酶体靶向E-cadherin:在上皮到间质转变过程中下调细胞-细胞粘附的独特机制。摩尔细胞生物25日:389 - 402。[Crossref]
- Figueroa A, Kotani H, Toda Y, Mazan-Mamczarz K, Mueller EC,等(2009)hakai在细胞增殖和肿瘤发生中的新作用。细胞杂志20: 3533 - 3542。[Crossref]
- Ordóñez-Morán P, Larriba MJ, Pálmer HG, Valero RA, Barbáchano A,等(2008)RhoA-ROCK和p38MAPK-MSK1介导维生素D对结肠癌细胞基因表达、表型和Wnt通路的影响。J细胞183: 697 - 710。[Crossref]
- Gozani O, Karuman P, Jones DR, Ivanov D, Cha J,等(2003)博士色谱相关蛋白ING2作为核磷酸肌苷受体的功能。细胞114: 99 - 111。[Crossref]
- Bua DJ, Martin GM, Binda O, Gozani O(2013)核磷脂酰肌醇-5-磷酸调节ING2在离散染色质靶上的稳定性,以响应DNA损伤。Sci代表3: 1 - 8。
- Le Jan S, Amy C, Cazes A, Monnot C, Lamande N et al.(2003)血管生成素样4是缺血和常规肾细胞癌中产生的促血管生成因子。是中草药162: 1521 - 1528。[Crossref]
- Young JA, Ting KK, Li J, Moller T, Dunn L,等(2013)miR-27的一般和ve -cadherin限制性miRNA拮抗剂对血管泄漏和缺血损伤恢复的调控作用。血122: 2911 - 2919。[Crossref]
- Semenza GL(2010)血管对缺氧和缺血的反应。动脉粥样硬化血栓血管生物学30: 648 - 652。[Crossref]
- 沈强,王颖,Kokovay E,林刚,庄思明,等(2008)成体SVZ干细胞位于血管生态位:生态位细胞与细胞相互作用的定量分析。细胞干细胞3: 289 - 300。[Crossref]
- Young CC, van der Harg JM, Lewis NJ, Brooks KJ, Buchan AM,等(2013)脑卒中后重组室管膜纤毛功能障碍和反应性星形细胞增多。大脑皮层23日:647 - 659。[Crossref]
- Zechariah A, ElAli A, Doeppner TR, Jin F, Hasan MR,等(2013)血管内皮生长因子促进脑毛细血管周细胞覆盖,改善继发局灶性脑缺血时的脑血流,并保留代谢半暗带。中风44: 1690 - 1697。[Crossref]
- Licht T, Dor-Wollman T, Ben-Zvi A, Rothe G, and Keshet E.(2015)血管成熟时间表决定了早产儿神经元损伤的易损性。中国投资125: 1319 - 1328。[Crossref]
- van Bruggen N, Thibodeaux H, Palmer JT, Lee WP, Fu L等(1999)VEGF拮抗减少小鼠脑缺血/再灌注损伤后水肿形成和组织损伤。中国投资104: 1613 - 1620。[Crossref]
- Fernández-Lopez D, Faustino J, Derugin N, Vexler ZS(2013)实验性局灶性动脉卒中对新生大鼠急性和慢性血管反应。Transl中风Res4: 179 - 188。
- Silbereis JC, Huang EJ, Back SA, Rowitch DH(2010)改进脑性瘫痪相关新生儿白质损伤动物模型。说模型动力机械3: 678 - 688。[Crossref]
- Dzietko M, Derugin N, Wendland MF, Vexler ZS, Ferriero DM(2013)延迟VEGF治疗增强新生鼠局灶性脑卒中后的血管生成和恢复。Transl中风Res4: 189 - 200。[Crossref]
- israel I, Costa RM, Xie CW, Silva AJ, Kosik KS,等(2004)神经元特异性蛋白delta-catenin的缺失导致严重的认知和突触功能障碍。咕咕叫杂志14: 1657 - 1663。[Crossref]
- Arikkath J, Peng IF, Ng YG, israel I, Liu X,等(2009)Delta-catenin在发育过程中调节脊柱和突触的形态发生和海马神经元的功能。J >29日:5435 - 5442。[Crossref]
- Pálmer HG, González-Sancho JM, Espada J, Berciano MT, Puig I, et .(2001)维生素D(3)通过诱导E-cadherin和抑制-catenin信号通路促进结肠癌细胞分化。J细胞154: 369 - 387。[Crossref]


