抽象的
缺血再灌注或创伤性脑损伤均可诱导脑内活性氧(ROS)的积累。ROS引起星形胶质细胞和神经元的氧化应激,氧化应激导致细胞器、蛋白质或脂质损伤。去除受损细胞的胞质成分是维持细胞内环境稳定不可缺少的。在大细胞自噬(以下简称自噬)中,降解缺陷蛋白和受损细胞器的过程,受损的细胞质成分是通过自噬体分离出来的。微管相关蛋白1轻链3B (LC3B)在自噬体形成中起重要作用,未偶联的LC3B (LC3B- i)向磷脂酰乙醇胺(PE)偶联的LC3B (LC3B- ii)的转化是自噬激活的指标。γ-氨基丁酸-a型受体相关蛋白样1 (GABARAPL1)是LC3B的一个类似物;它的功能还没有被完全理解。
在本研究中,我们证明了GABARAPL1 mRNA和蛋白的表达被过氧化氢(H2O2)在大鼠C6胶质瘤细胞中,并将Gabarapl1诱导h2O2伴随着从LC3B-I转化为LC3B-II的转化,表明形成自噬体。因此,GabaraPL1可以与由H引起的自噬过程相关联2O2.
关键字
自噬,反应性氧气物种,LC3,加巴帕
缩写
ROS:反应性氧气种类,H2O2:过氧化氢,LC3:微管相关蛋白1轻链,PE:磷脂酰乙醇胺,GABARAP: γ氨基丁酸-a型受体相关蛋白,CQ:氯喹
介绍
缺血再灌注或创伤性脑损伤导致中枢神经系统活性氧(ROS)的产生和积累[1,2]。ROS引起神经元和星形胶质细胞的氧化应激,氧化应激导致细胞胞质成分的损伤,如线粒体、蛋白质或脂质。细胞的存活和内稳态的维持依赖于受损细胞的胞质成分的去除。巨噬(Macroautophagy,以下简称自噬)是在包括氧化应激在内的各种胁迫下,缺陷蛋白和受损细胞器被分离和降解,生成新的蛋白和细胞器的过程[3-5]。
自噬经历了一系列的阶段,包括起始、延伸、闭合和成熟。启动自噬的一个重要过程涉及抑制哺乳动物(或机制)雷帕霉素靶点(mTOR), ROS抑制mTOR[6-9]。在自噬的伸长和闭合阶段,形成一种称为自噬小体的双层膜结构,以分离细胞的胞质成分。自噬小体的形成是由微管相关蛋白1轻链3 (LC3)亚家族成员和γ-氨基丁酸-a受体相关蛋白(GABARAP)亚家族成员介导的。在大鼠中,LC3亚家族包括LC3A和LC3B, GABARAP亚家族包括GABARAP、GABARAP-like 1 (GABARAPL1)和GABARAP-like 2 (GABARAPL2)[10-14]。这些成员与磷脂酰乙醇胺(PE)结合,而PE结合的这些成员形式在自噬体的形成中起着重要作用[11,13,15]。unconjugated-LC3B (LC3B- i)向PE-conjugated LC3B (LC3B- ii)的转化表明自噬体[13]的形成。此外,有研究报道GABARAPL1在受损线粒体[16]和缺氧[17]的降解中发挥重要作用。然而,包括过氧化氢(H2O2)仍然没有撇开。
星形胶质细胞在中枢神经系统中有几个重要的作用:为神经元提供营养、形成血脑屏障、维持细胞外离子平衡、包围和维持突触、参与神经元的信息加工[18-20]。因此,大脑的最佳功能依赖于自噬来保护星形胶质细胞和神经元免受氧化应激。
在这项研究中,我们检查了H的影响2O2大鼠C6胶质瘤细胞常被用作星形胶质细胞模型,表达LC3B和GABARAPL1 [21] mRNA。
材料和方法
材料
H2O2二磷酸氯喹从日本大阪Wako纯化学工业公司购买。Anti-GABARAPL1 (11010-1-AP)从Proteintech (Chicago, IL, USA)获得。抗肌动蛋白抗体来自Sigma(美国MO, St. Louis)。Anti-LC3 (8E10)和辣根过氧化物酶相关二抗来自MBL(日本名古屋)。
细胞培养
从日本大阪的JCRB细胞库中获得大鼠C6胶质瘤细胞,并在37°C、5% CO湿化条件下培养2Dulbecco的改性鹰培养基(DMEM)的气氛补充有10%FBS,100个单位/ ml青霉素和100μg/ ml链霉素。
实时定量聚合酶链反应
用NucleoSpin RNA (machery - nagel, Düren, Germany)纯化C6胶质瘤细胞的总RNA。逆转录酶反应进行使用PrimeScript II 1日1μg的RNA链cDNA合成装备(豆类生物,大津,日本)在20毫升卷42°C 60分钟。实时定量PCR进行与SYBR预混料交货Taq II(豆类生物,大津,日本)和LineGene (BioFlux、东京、日本)使用以下引物:LC3B, 5 ' -GAGCTTCGAACAAAGAGTGGA-3 '和5 ' -CTTCTCACCCTTGTATCGCTCTA-3 ';GABARAPL1, 5 ' -AGAAGGCTCCTAAAGCCAGG-3 '和5 ' -GTCCTCAGGTCTCAGGTGGA-3 ';羟甲基双烷合成酶(HMBS), 5 ' - TGGCGCAGCTACAGAGAAAG-3 '和5 ' - AGCTCATCCAGCTTCCGTAG-3 '。使用cDNA序列稀释生成相对标准曲线,以确定每个目标基因的扩增效率。将目的基因的表达量与每个样本的HMBS表达量进行比较。使用REST软件[22]计算相对表达量。
Western印迹分析
C6胶质瘤细胞被收集并溶解在改良的RIPA缓冲液(50 mM Tris-HCl, pH 7.4, 1% NP-40, 0.25%脱氧胆酸钠,150 mM NaCl, 1 mM EDTA, 2 mM氟磷酸二异丙酯,10 mg/mL的leptin, 10 mg/mL的抑肽酶,10 mg/mL的pepstatin)中。细胞裂解液离心,上清液中的蛋白质在十二烷基硫酸钠-聚丙烯酰胺凝胶中电泳,并转移到Immobilon P膜(Millipore, Billerica, MA)。用1%脱脂牛奶和0.1% Tween 20/PBS (PBS- t)封闭膜,并与一抗孵育。使用辣根过氧化物酶结合的二抗,免疫反应蛋白用ImmunoStar LD (Wako Pure Chemical Industries, Osaka, Japan)检测。
统计分析
多次观察的结果显示为平均值±扫描电镜。组间差异通过Student's进行分析t测试。P<0.05被认为是显著的。
结果
h的效果2O2对C6胶质瘤细胞LC3B和GABARAPL1 mRNA表达的影响
通过定量PCR评估C6胶质瘤细胞中LC3B和GABARAPL1的mRNA表达。用表明浓度的H处理C6胶质瘤细胞2O2,6小时后提取总RNA(图1A)。H未显著提高LC3B mRNA表达水平2O2.相反,在0.25 mM H时,GABARAPL1 mRNA的表达量增加到对照的3倍2O2而GABARAPL1 mRNA的表达量在0.5 mM H时达到最大值(为对照的5倍)2O2.该表达保持在1 mM H2O2.我们还研究了1mm H的影响2O2LC3B和GABARAPL1在不同时间点的mRNA表达(图1B)。H未显著提高LC3B mRNA表达水平2O2相比之下,GABARAPL1 mRNA表达量在4 h时增加到对照的2.5倍,在6 h时GABARAPL1 mRNA表达量增加达到最大值(是对照的5倍),该表达维持到8 h。
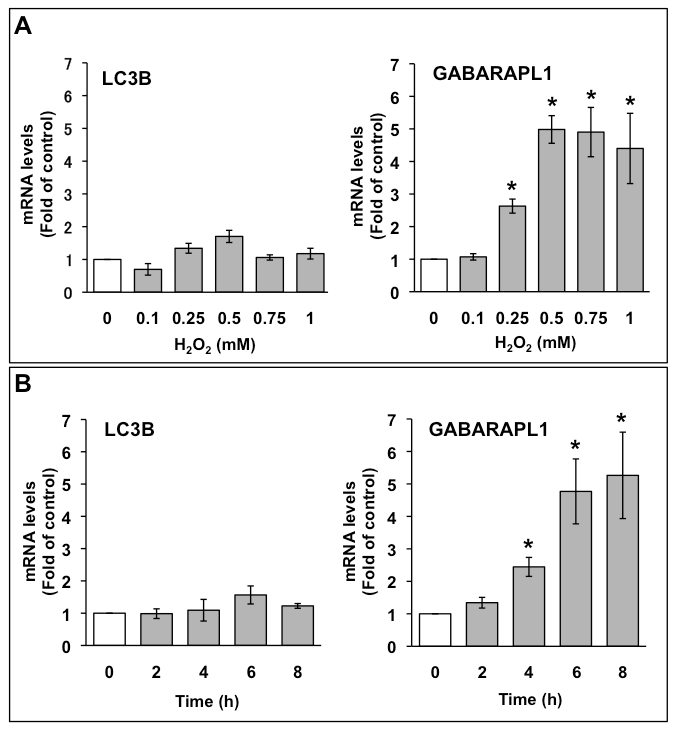
图1所示。A. H2O2对C6胶质瘤细胞LC3B和GABARAPL1 mRNA表达的剂量依赖性影响用相应浓度的H2O2处理C6胶质瘤细胞6 h。(B) H2O2处理C6胶质瘤细胞LC3B和GABARAPL1 mRNA表达的时间历程。用1mm H2O2处理C6胶质瘤细胞。将LC3B、GABARAPL1 mRNA表达水平与HMBS mRNA表达水平进行比较,以HMBS mRNA作为内对照。结果是治疗水平与对照组的比值(平均值±扫描电镜,n = 3)。P<0.05被认为有统计学意义。
GABARAPL1-I,而不是GABARAPL1-II,在H2O2在C6胶质瘤细胞中
用50µM氯喹(CQ)处理C6胶质瘤细胞16小时。CQ是溶酶体降解的抑制剂,可抑制pe标记的LC3B (LC3B- ii)在溶酶体中的降解。在SDS-PAGE上,LC3B- ii和pe标记的GABARAPL1 (GABARAPL1- ii)分别比未标记的LC3B (LC3B- i)和未标记的GABARAPL1 (GABARAPL1- i)迁移更快。在CQ不存在和存在时均可检测到LC3B-II。虽然在CQ不存在的C6胶质瘤细胞中检测到GABARAPL1-I,但仅在CQ存在的C6胶质瘤细胞中检测到GABARAPL1-II(图2A)。接下来,我们用1mm H检测LC3B和GABARAPL1的表达2O2.因为LC3B从LC3B- i到LC3B- ii的转化是自噬体形成的关键步骤,表明自噬体数量的增加,1 mM H的作用2O2在不同时间点检测LC3B-II水平,持续8 h(图2B)。1 mM H时LC3B-II水平约为对照的2倍2O24小时后,保持在此水平。1 mM H时,LC3B总表达水平轻微但显著升高2O2.虽然没有检测到Gabarapl1-II,但是2O2-处理C6胶质瘤细胞,GABARAPL1-I表达水平增加到对照组的两倍2O2之后GABARAPL1-I表达水平维持在该水平(图2C)。

图2。H2O2处理后GABARAPL1-I表达增加。C6胶质瘤细胞用50µM chloroquin (CQ)处理16小时。细胞裂解液用抗lc3抗体或抗gabarapl1抗体(A)进行Western blot分析。C6胶质瘤细胞用1 mM H2O2处理指定的时间。细胞裂解液用抗lc3抗体(B)或抗gabarapl1抗体(C)进行Western blot分析。以肌动蛋白表达水平作为内对照。结果是治疗水平与对照组的比值(平均值±扫描电镜,n = 3)。P<0.05被认为有统计学意义。
GABARAPL1通过H2O2是否受C6胶质瘤细胞中溶酶体降解抑制的影响
LC3B-II存在于内部和外部自血糖体膜上,并且在成熟到自糖体后,内自血管体膜上的LC3B-II通过溶酶体降解。相反,外自血管体膜上的LC3B-II被齐平并重复使用以形成自噬体[23]。
LC3B-II水平的增加表明自糖蛋白酶中LC3B-II的降低降低增加。因此,LC3B-II水平与表明自噬活性的自噬助体无关,尽管它们与自噬复印件的数量有关。如果LC3B-II的水平由H2O2有溶酶体降解抑制剂的治疗比没有溶酶体降解抑制剂的治疗高,这表明自噬通量[24]增加。因此,我们检测了1mm H的效果2O2在8小时内使用50μmcq的自噬助焊剂。H通过H增加LC3B-II水平2O2在CQ存在时升高(图3A)。基于这些结果,H2O2治疗可能是由于自噬通量的增强。而H2O2在CQ存在的情况下受影响不大(图3A)。此外,我们还检测了CQ对GABARAPL1表达的影响。GABARAPL1-II在CQ存在8 h时未检测到2O2在CQ的存在下没有太大改变(图3b)。

图3。GABARAPL1-I在H2O2治疗不受CQ的影响。用1mm H处理C6胶质瘤细胞2O2用抗lc3b抗体(A)或抗gabarapl1抗体(B)对细胞裂解液进行Western blot分析。以肌动蛋白表达水平作为内对照。结果是治疗水平与对照组的比值(平均值±扫描电镜,n = 3)。P<0.05被认为有统计学意义。
讨论
2021年版权燕麦。所有权利reserv
在缺血再灌注或外伤性脑损伤的情况下,中枢神经系统会产生并积累ROS[1,2]。超氧化物主要在ROS中产生,然后迅速转化为H2O2通过减少。此外,H2O2比其他活性氧更稳定。因此,H2O2通过氧化应激或作为细胞内信号信使影响细胞的生理反应。H2O2通过血浆膜上的水蛋白转化为细胞[25],和h2O2增加了H2O2并可导致线粒体损伤[26,27]。此外,H2O2也会在细胞内通过饥饿和线粒体损伤产生,导致氧化应激。氧化应激与自噬[3]高度相关。H2O2抑制mTOR,启动自噬的关键调控因子[6-9]。此外,H2O2抑制ATG4, ATG4释放LC3-II,导致LC3B-II水平升高,自噬体[28]形成。自噬可以保护脑细胞免受氧化应激的损伤。虽然自噬体的形成和LC3B-II水平是由H2O2[9,30], H2O2在星形胶质细胞中未被阐明。因此,我们通过引入H2O2.H mRNA表达水平的H mRNA表达水平大大增加2O2.我们证实H2O2而H2O2自噬通量增加,因为CQ抑制溶酶体降解增强了H2O2.相比之下,虽然GABARAPL1- i (GABARAPL1的非共轭形式)在C6胶质瘤细胞中表达增加,但未检测到GABARAPL1- ii (pe -共轭形式的GABARAPL1)2O2.此外,H诱导的GabaraPL1-I表达的增加2O2受CQ的影响不大。因此,这表明GABARAPL1-I没有被吞噬到自噬体中,也没有在自溶酶体中被降解。GABARAPL1-I可能在自噬体外膜或外膜起作用。LC3亚家族和GABARAP亚家族成员参与了自噬小体的形成[11,13,15]以及自噬小体与溶酶体的融合[31,32]。有报道称LC3B在自噬体形成早期起作用,GABARAPL2在自噬体闭合阶段[15]起作用。相反,LC3亚家族和GABARAP亚家族成员结合具有LC3相互作用区域(LIR)的自噬适配蛋白[13,33]。LIR在LC3亚家族和GABARAP亚家族成员中结合W/FxxL基序,也称为atg8 -家族相互作用基序(AIM)[13,34,35]。然而,LC3亚家族和GABARAP亚家族的每个成员都可以与不同的蛋白相互作用,在自噬中具有离散的功能。此外,研究表明LC3亚家族和GABARAP亚家族成员可以以aim依赖和非依赖的方式结合其靶蛋白,并且这些成员可以与不止一个蛋白[36]相互作用。由于GABARAPL1可以与GABARAPL2[36]结合,它可能通过GABARAPL2关闭自噬小体,或者通过结合GABARAPL2使自噬小体与溶酶体融合。 Conversely, many proteins that can bind to GABARAPL1 can also bind to the other members of LC3 subfamily and GABARAP subfamily [36]. Therefore, as the unconjugated form of GABARAPL1 (GABARAPL1-I) is not able to associate with autophagosomal membranes, it is possible that increased GABARAPL1-I may function to bind to its target proteins for protecting the degradation of these proteins by blocking the binding of the PE-conjugated form of the other LC3 subfamily and GABARAP subfamily members, which can bring these proteins into autophagosomes. This may lead to the selectivity of the degradation of autophagic cargoes by either PE-conjugated or -unconjugated form of LC3 subfamily and GABARAP subfamily members.
综上所述,我们证明了GABARAPL1 mRNA在H2O2在C6胶质瘤细胞中。我们还表明,H-I水平的增加2O2.H2O2GABARAPL1的功能有待进一步研究。
总结
活性氧会引起细胞器或蛋白质的损伤。受损的细胞胞质成分通过自噬体分离并通过自噬过程降解。从未偶联LC3B (LC3B- i)到磷脂酰乙醇胺偶联LC3B (LC3B- ii)的转化是自噬激活的指标。γ-氨基丁酸-a型受体相关蛋白样1 (GABARAPL1)是LC3B的一个类似物;它的功能还没有被完全理解。我们证实了GABARAPL1 mRNA和蛋白的表达被过氧化氢(H2O2)在大鼠C6胶质瘤细胞中,并将Gabarapl1诱导h2O2伴随着从LC3B-I转换到LC3B-II的转换。Gabarapl1可能与H引起的自噬过程相关联2O2.
利益冲突
作者声明没有利益冲突。
参考
- Chen H, Yoshioka H, Kim GS, Jung JE, Okami N, et al.(2011)缺血性脑损伤的氧化应激:细胞死亡机制和潜在的神经保护分子靶点。Antioxid氧化还原信号14: 1505 - 1517。[Crossref]
- Werner C, Engelhard K(2007)创伤性脑损伤的病理生理学。Br J Anaesth99: 4 - 9日。[Crossref]
- 自噬与氧化应激之间的相互作用。细胞死亡是不同的22:377-388。[Crossref]
- Kiriyama Y, Kino K, Nochi H(2015)自噬与氨基酸及其代谢产物。中国食物减轻金属底座2: 151 - 155。
- 自噬与综合应激反应的关系。摩尔细胞40:280-293。[Crossref]
- Alexander A, Cai SL, Kim J, Nanez A, Sahin M,等(2010)细胞质中ATM信号对TSC2调控mTORC1应答ROS。美国国立科学院科学研究所107: 4153 - 4158。[Crossref]
- Byun YJ,Kim Sk,Kim Ym,Chae Gt,Jeong Sw,等。(2009)过氧化氢通过BNIP3介导的MTOR途径抑制C6胶质瘤细胞中的自噬细胞死亡。>列托人461: 131 - 135。[Crossref]
- 刘L,Wise Dr,Diehl Ja,Simon MC(2008)缺氧反应性氧物种调节综合应力响应和细胞存活。J临床生物化学283: 31153 - 31162。[Crossref]
- 张H,孔X,康J,Su J,Li Y等。(2009)氧化应激在人胶质瘤U251细胞中诱导平行的自噬和线粒体功能障碍。Toxicol Sci110: 376 - 388。[Crossref]
- Kabeya Y,Mizushima N,Ueno T,Yamamoto A,Kirisako T,等。(2000)LC3,酵母APG8P的哺乳动物同源物,在加工后局部局部化在自噬体膜中。EMBO J19:5720-5728。[Crossref]
- Kabeya Y, Mizushima N, Yamamoto A, Oshitani-Okamoto S, Ohsumi Y, et al. (2004) LC3, GABARAP和GATE16定位于自噬体膜依赖于form-II的形成。J细胞科学117:2805-2812。[Crossref]
- Tanida I, Komatsu M, Ueno T, Kominami E (2003) GATE-16和GABARAP是Apg7和Apg3介导的可信修饰词。Biochem Biophys Res Communic300:637-644。[Crossref]
- 野生p,mcewan dg,迪克斯i(2014)LC3互乱一目了然。J细胞科学127: 3 - 9。[Crossref]
- Xin Y, Yu L, Chen Z, Zheng L, Fu Q, et al. (2001) GABA(A)受体相关蛋白3个人类和2个小鼠同源基因的克隆、表达模式和染色体定位。基因组学74: 408 - 413。[Crossref]
- Weidberg H,Shvets E,Shpilka T,Shimron F,Shinder v等。(2010)LC3和Gate-16 / Gabarap Subcamilies既是必需的,但在自噬生物生物发生时也不同。EMBO J29日:1792 - 1802。[Crossref]
- noak I, Kirkin V, McEwan DG, Zhang J, Wild P, et al. (2010) Nix是一种线粒体清除的选择性自噬受体。Embo Rep.11: 45-51。[Crossref]
- Keulers TG, Schaaf MB, pepeters HJ, Savelkouls KG, Vooijs MA, et al.(2015)缺氧期间EGFR膜表达增加需要GABARAPL1。Radiother杂志116: 417 - 422。[Crossref]
- Allaman I,Bélangerm,Magistretti PJ(2011)星形胶质细胞 - 神经元代谢关系:好转,而且差了。趋势>34: 76 - 87。[Crossref]
- (2004)血脑屏障:概述:结构、调节和临床意义。一般人说16: 1-13。[Crossref]
- Parpura V,Heneka Mt,蒙大拿州v,oliet sh,schousboe a等。(2012)(Patho)生理学中的胶质细胞。J Neurochem121:速率。[Crossref]
- Kiriyama Y, Ozaki A, Kino K, Nochi H (2015) CCCP对C6胶质瘤细胞中GABARAPL2表达的影响。中国摩尔地中海2:162-165。
- Pfaffl MW,Horgan GW,Dempfle L(2002)相对表达软件工具(REST)用于群体的比较和相对表达的统计分析导致实时PCR。核酸Res30: e36。[Crossref]
- Klionsky DJ, Schulman BA2(2014)特异性泛素样蛋白对巨噬细胞的动态调控。Nat Struct Mol Biol21日:336 - 345。[Crossref]
- Klionsky DJ, Abdelmohsen K, Abe A, Abedin MJ, Abeliovich H, et al.(2016)自噬检测方法的使用和解释指南(第3版)。自噬12:1-222。[Crossref]
- Bienert GP和Chaumont F(2014)水通道蛋白促进过氧化氢跨膜扩散。Biochim Biophys学报1840: 1596 - 1604。[Crossref]
- Kudin AP, Bimpong-Buta NY, Vielhaber S, Elger CE, Kunz WS(2004)分离的脑线粒体中超氧化物产生位点的特征。J临床生物化学279: 4127 - 4135。[Crossref]
- Lin MT, Beal MF(2006)线粒体功能障碍与神经退行性疾病的氧化应激。自然443: 787 - 795。[Crossref]
- Scherz-Shouval R, Shvets E, Fass E, Shorer H, Gil L, et al.(2007)活性氧是自噬所必需的,并特别调节Atg4的活性。EMBO J26:1749-1760。[Crossref]
- 自噬在神经退行性疾病中的作用[j]。氧化还原生物2: 82 - 90。[Crossref]
- Zhang J, Kim J, Alexander A, Cai S, Tripathi DN,等。Nat细胞生物15: 1186 - 1196。[Crossref]
- Wang H, Sun HQ, Zhu X, Zhang L, Albanesi J2, et al. (2015) GABARAPs调节pi4p依赖的自噬体:溶酶体融合。美国国立科学院科学研究所112: 7015 - 7020。[Crossref]
- Wilkinson DS, Jariwala JS, Anderson E2, Mitra K3, Meisenhelder J4, et al.(2015)通过Hippo激酶STK3/STK4磷酸化LC3是自噬的关键。摩尔细胞57: 55 - 68。[Crossref]
- Johansen T, Lamark T(2011)自噬适配器蛋白介导的选择性自噬。自噬7: 279 - 296。[Crossref]
- Birgisdottir ÅB, Lamark T, Johansen T (2013) LIR基序——选择性自噬的关键。J细胞科学126: 3237 - 3247。[Crossref]
- Rozenknop A, Rogov VV, Rogova NY, Löhr F, Güntert P, et al. (2011) GABARAPL-1与NBR1 LIR基序相互作用的表征。J杂志410: 477 - 487。[Crossref]
- Behrends C, Sowa ME, Gygi SP, Harper JW(2010)人体自噬系统的网络组织。自然466: 68 - 76。[Crossref]



