摘要
环氧合酶-2 (COX-2)在结直肠癌的发生中起着至关重要的作用。对COX-2基因变异(-765G>C, -1195G>A和8473T>C)与结直肠癌(CRC)易感性之间关系的研究积累得出了相互矛盾的结果。为了探索潜在的相关性,我们基于合格的病例对照流行病学研究进行了meta分析。筛选Medline、Embase和CNKI数据库中关注COX-2多态性与CRC风险关系的合格论文。计算合并比值比(OR)和95%置信区间(95% CI)来估计这种关联的显著性。本荟萃分析共纳入了12项-765G>C研究、8项COX-2 -1195G>A研究和7项8473T>C研究。总体而言,-1195 A等位基因携带者的CRC发病率高于G等位基因携带者(OR=1.227, 95% CI=1.060-1.419,P或= 0.006)。样本量≥500的亚组分析结果一致(OR=1.232, 95% CI=1.064-1.426,P或= 0.005)。对于-765C等位基因,仅在杂合子和显性模型下的亚洲后裔中观察到较高的CRC风险(OR=1.477, 95% CI=1.148-1.898,P或= 0.002;Or =1.451, 95% ci =1.146- 1.747,P或分别为= 0.001)。相比之下,8473T>C多态性与CRC易感性之间在所有4种遗传模型下均未定义关联。考虑到基因背景和环境因素对结直肠癌的交互影响,-765GG基因型吸烟者发生结直肠癌的风险比对照组高1.761倍。总之,-1195G>A多态性在世界范围内是CRC易感性的因素,而-765G>C仅在亚洲人群中是潜在的CRC危险因素。-765GG基因型吸烟者更容易发生结直肠癌。考虑到基因-基因和基因-环境相互作用的大样本、匹配良好的研究正在等待验证这些发现。
关键字
基因变异,COX-2,结直肠癌,荟萃分析
简介
结直肠癌(CRC)是全球男性和女性癌症死亡的主要常见原因[1]。2013年,估计有15万例CRC新诊断病例,其中三分之一以上的患者预计将死于这种疾病,占所有癌症相关死亡的9%[2]。虽然结肠直肠癌发生的分子机制尚不清楚,但炎症与结直肠癌发生之间的联系已得到广泛认识。
作为一种促炎酶,环氧合酶-2 (COX-2)对促炎和有丝分裂刺激的反应显著增强,导致花生四烯酸[4]多余地合成前列腺素。COX-2通过影响细胞增殖、细胞凋亡、血管生成和免疫反应,被认为在致癌过程中起着关键作用[5,6]。值得注意的是,近85%的CRC患者表现出COX-2的高表达,这与CRC患者的晚期和生存期较差有关[7,8]。巧合的是,据广泛报道,被称为COX-2抑制剂的非甾体抗炎药(NSAIDs)是最有希望预防结直肠癌的药物之一。易感人群接受非甾体抗炎药治疗可使CRC发生率降低约60%[10]。以上信息均支持COX-2对结直肠癌的发育有重要影响,可能是结直肠癌预防的分子靶点[11]。
众所周知,COX-2酶的功能依赖于COX-2的表达和活性。COX-2的功能性遗传变异可能改变COX-2酶的表达和活性,从而影响个体对CRC发生的易感性。到目前为止,在定位于1q25.2-q25.3的COX-2基因中已经发现了许多功能性多态性。其中-765G>C、-1195G>A和8473T>C是CRC中研究最广泛的多态性。第一项关于-765G>C、-1195G>A和8473T>C多态性对发生CRC风险影响的研究分别发表于2001年[12]、2004年[13]和2006年[14]。此后出现了大量的研究[15,16-27]。然而,这些研究结果是有争议的。以COX-2 -1195G>A多态性为例,Siezen的研究结果表明COX-2 -1195G>A与CRC发生[15]的风险无关。相比之下,Tan等人的另一项研究表明,它们之间存在正相关。为了阐明这些不确定的发现,我们进行了当前的meta分析,定量评估上述三种多态性对CRC易感性的潜在影响。 Interactive influence between environmental factors and gene polymorphisms on CRC risk would be also estimated if there was enough data for OR and 95%CI calculation.
方法
搜寻符合条件研究的策略
我们筛选了Medline、Embase和CNKI(中国国家知识基础设施)数据库中COX-2多态性与CRC易感性之间关系的相关研究。关键词如下:“环氧合酶-2”或“COX-2”或“前列腺素合成酶2”或“PTGS2”,“结直肠癌”或“结直肠腺癌”,“多态性”或“变异”或“基因型”或“单核苷酸多态性”或“SNP”,以及“风险”或“易感”(最近一次搜索更新于2013年8月20日)。检索到的综述或荟萃分析中相应的参考文献被手动筛选以获得额外的合格研究。本荟萃分析不考虑语言限制。采用以下纳入标准筛选符合条件的研究:(1)病例对照研究;(2)探讨COX-2基因多态性对结直肠癌发生的可能影响;(3)重点研究COX-2的3个常见多态性:-765G>C、-1195G>A和8473T>C;(4)经组织病理证实为结直肠癌;(5)有足够的数据计算比值比(OR)和95%置信区间(95% ci)。此外,如果纳入的研究包含重叠或相同的主题,将选择最新的、样本量最大的研究。
数据提取
两位作者(Pan Y和Gao S)从所有合格的论文中独立审阅并检索了有用的信息。从每项研究中提取以下参数:发表年份、第一作者、国家、种族(亚裔、白种人或混合血统)、对照来源(基于医院或基于人群)、病例和对照总数、基因分型方法和调整后的潜在混杂变量(表1)。通过所有作者之间的讨论来解决差异,以达成共识。
表S1。本综合荟萃分析中16项合格研究的特征
PCC,以人群为基础的病例对照;肝细胞癌,医院病例对照;PCR-CTPP,聚合酶链反应,对抗双对引物;PCR-RFLP,聚合酶链反应-限制性片段长度多态性分析;NA,不可用
第一作者(年) |
国家 |
种族 |
控制的起源 |
情况下 |
控制 |
基因分型方法 |
调整了潜在的混杂变量 |
Hamajima (2001) [12] |
日本 |
亚洲 |
肝细胞癌 |
148 |
241 |
PCR-CTPP |
NA |
考克斯(2004)[13] |
西班牙 |
高加索人 |
肝细胞癌 |
290 |
272 |
TaqMan化验 |
调整了年龄,性别,吸烟状态持续时间和BMI |
Koh (2004) [14] |
新加坡 |
亚洲 |
PCC |
310 |
1177 |
TaqMan化验 |
根据酒精消费调整;但对年龄、性别、身体质量指数和吸烟状况没有影响 |
Siezen (2006) [15] |
荷兰 |
高加索人 |
PCC |
682 |
1142 |
PCR-RFLP |
根据年龄和性别调整 |
谭(2007)[16] |
中国 |
亚洲 |
肝细胞癌 |
1000 |
1300 |
PCR-RFLP |
根据年龄和性别调整 |
兴(2008)[17] |
中国 |
亚洲 |
肝细胞癌 |
137 |
199 |
PCR-RFLP |
根据年龄,性别,吸烟状况,饮酒时间和BMI进行调整 |
伊格莱西亚斯(2009)[18] |
西班牙 |
高加索人 |
肝细胞癌 |
284 |
123 |
PCR-RFLP |
根据年龄和性别调整 |
汤普森(2009)[19] |
美国 |
高加索人 |
PCC |
421 |
480 |
TaqMan化验 |
根据吸烟状况调整;但对年龄、性别、身体质量指数和饮酒状况没有影响 |
霍夫(2009)[20] |
荷兰 |
高加索人 |
肝细胞癌 |
326 |
369 |
PCR-RFLP |
根据年龄和性别调整 |
安徒生(2009)[21] |
丹麦 |
高加索人 |
PCC |
359 |
765 |
TaqMan化验 |
调整的性别 |
佩雷拉(2010)[22] |
葡萄牙 |
高加索人 |
肝细胞癌 |
117 |
256 |
PCR-RFLP或rt - pcr |
根据性别、BMI和吸烟状况进行了调整;但不是因为年龄 |
王(2012)[23] |
美国 |
高加索人 |
肝细胞癌 |
305 |
359 |
TaqMan化验 |
根据年龄和性别调整 |
Daraei (2012) [24] |
伊朗 |
亚洲 |
PCC |
110 |
120 |
PCR-RFLP |
根据年龄和性别,身体质量指数和吸烟状况进行调整 |
张(2012)[25] |
中国 |
亚洲 |
肝细胞癌 |
343 |
340 |
PCR-RFLP |
根据年龄、性别、吸烟状况和饮酒状况进行调整 |
罗斯(2013)[26] |
澳大利亚、西班牙语 |
高加索人 |
肝细胞癌 |
183 |
223 |
PIRA试验,TaqMan试验 |
NA |
李(2013)[27] |
中国 |
亚洲 |
肝细胞癌 |
451 |
629 |
PCR-RFLP |
调整的性别;但对年龄和BMI没有影响 |
统计分析
在杂合子模型、变异纯合子模型、显性模型和隐性模型4种遗传模型下,通过粗OR和95% CI估计COX-2多态性对个体发生结直肠癌差异易感性的影响。为了消除混杂因素的潜在影响,采用种族、样本量和对照来源等变量进行分层分析。
基于卡方的q检验和我2采用检验方法评价纳入研究的异质性[28,29]。如果p>0.05,采用固定效应模型[30]进行meta分析,否则采用随机效应模型[31]。在研究间异质性较高的情况下,建议进行敏感性分析以确认结论的一致性。发表偏倚方面,基本采用漏斗图、Begg秩相关法[32]和Egger加权回归法[33]。
各单项研究对照组基因型分布的HWE采用Pearson拟合优度卡方检验进行估计。采用STATA软件(12.0版本,STATA Corporation, USA)进行meta分析。P<0.05为差异显著。
结果
合资格研究的特点
在本荟萃分析中,我们使用荟萃分析方法定量评估了COX-2中三种常见遗传变异对CRC易感性的影响。总之,本研究最终纳入了12项-765G>C[12-14,16-24]、8项-1195G>A[15,16,19-22,25,27]和7项8473T>C[13,15,19,21,25,26]多态性研究,包括5466例CRC病例和7995例对照。研究选择流程图如图1所示。表1和表2列出16份合格出版物的详细资料。这三个多态性要么位于COX-2基因的启动子上,要么位于COX-2基因的3 ' -UTR上(图2)。在这些研究中,有7个研究来自亚洲人群,9个研究来自高加索族裔。关于对照来源,9项研究是基于医院的病例对照研究,其余是基于人群的病例对照研究。从所有研究对象的外周血中提取DNA,采用PCR- RFLP、PCR- ctpp或TaqMan法对COX-2的多态性进行基因分型。

图1所示。当前元分析的流程图

图2。人类COX-2基因的基因组结构由10个外显子和9个内含子组成。-1195G>A多态性和-765G>C多态性位于启动子区,8473T>C多态性位于COX-2的3'UTR区。
在当前的meta分析中,很明显,不同的多态性,-1195G>C, -765G>C和8473T>C,在可变遗传模型下对CRC的发展有不同的贡献。此外,诸如种族、对照来源、样本量、吸烟、非甾体抗炎药等环境因素等混杂因素也会影响这种关联的结果。因此,对每个COX-2多态性进行组合分析和亚组分析。
-1195G>A与CRC风险相关性的meta分析
8项-1195G>A的研究[15,16,19-22,25,27],共3648例和5219例对照,评估了COX-2 -1195G>A与CRC易感性的相关性。总体而言,COX-2基因变体-1195 a等位基因显著增加了CRC发生的风险(AGvs。Gg, or =1.227, 95% ci = 1.060-1.419,Ph= 0.085,P或= 0.006)。(图3,表S3)众所周知,大的样本量可以增加这种关系的统计力。有趣的是,我们发现在两种遗传模型下,在样本量≥500的亚组中,A等位基因与CRC易感性相关(AG vs. GG, OR=1.232, 95% CI=1.064-1.426,Ph= 0.056,P或= 0.005;AA vs. AG+GG, OR=1.162, 95% CI=1.006-1.342,Ph= 0.042,P或= 0.041)。(图4 A-B,表S3,)。此外,当排除两项偏离HWE的研究时,结果保持一致。敏感性分析证实-1195G>A多态性的所有结果都是稳定的。
表S2。各研究中特异性COX-2多态性和HWE的总结
HWE, Hardy-Weinberg平衡通过拟合优度x计算2以及。
作者 |
-765 g > C |
HWE |
-1195 g > |
HWE |
8473 t > C |
HWE |
GG |
CG |
CC |
GG |
AG) |
AA |
TT |
CT |
CC |
Hamajima [12] |
140/230 |
8/11 |
0/0 |
0.72 |
|
|
|
|
|
|
|
|
考克斯[13] |
150/170 |
59/77 |
11/10 |
0.73 |
|
|
|
|
140/126 |
121/120 |
29/25 |
0.64 |
Koh [14] |
273/1067 |
37/110 |
NA |
|
|
|
|
|
|
|
|
Siezen [15] |
|
|
|
|
29/61 |
191/354 |
410/665 |
0.13 |
313/529 |
254/444 |
75/108 |
0.3 |
谭[16] |
919/1237 |
81/63 |
0/0 |
0.37 |
178/300 |
502/692 |
320/308 |
0.02 |
|
|
|
|
邢[17] |
119/169 |
17/29 |
1/1 |
0.84 |
|
|
|
|
|
|
|
|
伊格莱西亚斯[18] |
172/76 |
99/43 |
13/4 |
0.48 |
|
|
|
|
|
|
|
|
汤普森[19] |
291/343 |
119/121 |
11/15 |
0.16 |
9/15 |
138/168 |
275/297 |
0.13 |
176/216 |
189/199 |
56/65 |
0.08 |
霍夫[20] |
241/249 |
75/112 |
10/8 |
0.17 |
12/13 |
101/124 |
213/232 |
0.47 |
|
|
|
|
安徒生[21] |
267/566 |
83/186 |
9/13 |
0.14 |
13/25 |
116/258 |
230/482 |
0.18 |
147/315 |
178/355 |
34/95 |
0.74 |
佩雷拉[22] |
77/166 |
38/83 |
2/7 |
0.19 |
4/6 |
43/73 |
70/177 |
0.63 |
54/118 |
51/114 |
10/24 |
0.64 |
王[23] |
207/238 |
87/111 |
11/10 |
0.18 |
|
|
|
|
|
|
|
|
Daraei [24] |
38/53 |
67/58 |
5/9 |
0.32 |
|
|
|
|
|
|
|
|
张[25] |
|
|
|
|
50/94 |
216/184 |
77/62 |
0.09 |
218/202 |
105/115 |
20/23 |
0.24 |
罗斯[26] |
|
|
|
|
|
|
|
|
74/107 |
93/83 |
16/33 |
0.01 |
李[27] |
|
|
|
|
87/114 |
248/336 |
116/179 |
0.05 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

图3。COX-2 -1195G>多态性与CRC易感性相关的森林图:AG vs. GG。
表3:COX-2三种常见多态性与CRC易感性之间的OR和P值总结
口服补液盐,优势比;cox - 2, cyclooxygenase-2;CRC,结直肠癌;CI,置信区间;Ht,杂合子;WT Ho,宽型纯合子;VR Ho,变异纯合子;Ph,PQ检验的异质性值。异质性检验P值<0.05时,采用随机效应模型;否则,采用固定模型。P或,P池或的值
遗传模型 |
|
Htvs。WT何 |
VR何vs。WT何 |
占主导地位的模式 |
隐性的模型 |
-765 g > C |
n |
CG / GG |
CC / GG |
CG + CC / GG |
CC / CG + GG |
|
|
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
总计 |
12 |
1.035 (0.871, 1.231) |
0.034 |
0.696 |
1.144 (0.811, 1.613) |
0.666 |
0.442 |
1.012 (0.881, 1.162) |
1.000 |
0.871 |
1.118 (0.801, 1.559) |
0.902 |
0.513 |
种族 |
高加索人 |
6 |
0.938 (0.811, 1.086) |
0.380 |
0.392 |
1.148 (0.778, 1.695) |
0.898 |
0.486 |
0.965 (0.831, 1.101) |
0.540 |
0.534 |
1.168 (0.794, 1.719) |
0.850 |
0.429 |
亚洲 |
5 |
1.477 (1.148, 1.898) |
0.242 |
0.002 |
0.847 (0.289, 2.479) |
0.694 |
0.762 |
1.415 (1.146, 1.747) |
0.387 |
0.001 |
0.665 (0.236, 1.874) |
0.553 |
0.440 |
混合 |
1 |
0.901 (0.643, 1.262) |
- |
0.545 |
1.265 (0.526, 3.038) |
- |
0.599 |
0.931 (0.673, 1.289) |
- |
0.667 |
1.306 (0.547, 3.118) |
- |
0.548 |
源的控制 |
PB |
4 |
1.107 (0.911, 1.345) |
0.224 |
0.307 |
0.870 (0.512, 1.479) |
0.248 |
0.608 |
0.987 (0.791, 1.232) |
0.935 |
0.908 |
0.941 (0.562, 1.575) |
0.394 |
0.817 |
乙肝 |
8 |
1.003 (0.758, 1.326) |
0.018 |
0.986 |
1.404 (0.827, 2.384) |
0.859 |
0.209 |
1.026 (0.840, 1.253) |
1.000 |
0.801 |
1.254 (0.753, 2.089) |
0.924 |
0.384 |
样本大小 |
≥500 |
7 |
1.009 (0.789, 1.289) |
0.006 |
0.946 |
1.187 (0.806, 1.750) |
0.923 |
0.385 |
1.061 (0.867, 1.299) |
0.016 |
0.563 |
1.216 (0.827, 1.787) |
0.866 |
0.320 |
< 500 |
5 |
1.088 (0.851, 1.391) |
0.566 |
0.500 |
0.970 (0.491, 1.918) |
0.808 |
0.931 |
1.078 (0.848, 1.370) |
0.678 |
0.540 |
0.872 (0.449, 1.693) |
0.682 |
0.685 |
-1195 g > |
n |
AG / GG |
AA / GG |
AG) + AA / GG |
AA / AG) + GG |
|
|
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
总计 |
8 |
1.227 (1.060, 1.419) |
0.085 |
0.006 |
1.287 (0.948, 1.749) |
0.009 |
0.106 |
1.255 (0.976, 1.613) |
0.030 |
0.077 |
1.109 (0.946, 1.301) |
0.008 |
0.203 |
种族 |
高加索人 |
5 |
1.048 (0.762, 1.442) |
0.910 |
0.771 |
1.145 (0.840, 1.560) |
0.686 |
0.392 |
1.112 (0.818, 1.510) |
0.781 |
0.498 |
1.083 (0.959, 1.223) |
0.269 |
0.199 |
亚洲 |
3. |
1.350 (0.902, 2.020) |
0.006 |
0.145 |
1.499 (0.871, 2.581) |
0.001 |
0.144 |
1.402 (0.917, 2.143) |
0.002 |
0.118 |
1.203 (0.837, 1.729) |
0.004 |
0.319 |
控制源 |
PB |
3. |
1.096 (0.766, 1.568) |
0.700 |
0.616 |
1.227 (0.866, 1.737) |
0.599 |
0.250 |
1.329 (1.141, 1.548) |
0.625 |
0.343 |
1.126 (0.981, 1.294) |
0.809 |
0.092 |
乙肝 |
5 |
1.253 (0.898, 1.750) |
0.023 |
0.185 |
1.297 (0.823, 2.042) |
0.002 |
0.262 |
1.274 (0.889, 1.826) |
0.007 |
0.187 |
1.077 (0.809, 1.435) |
0.001 |
0.611 |
样本大小 |
≥500 |
7 |
1.232 (1.064, 1.426) |
0.056 |
0.005 |
1.336 (0.980, 1.821) |
0.009 |
0.067 |
1.281 (0.991, 1.658) |
0.024 |
0.059 |
1.162 (1.006, 1.342) |
0.042 |
0.041 |
< 500 |
1 |
0.884 (0.236, 3.308) |
- |
0.854 |
0.593 (0.162, 2.166) |
- |
0.429 |
0.678 (0.188, 2.450) |
- |
0.553 |
0.665 (0.422, 1.048) |
- |
0.079 |
8473 t > C |
n |
CT / TT |
|
|
CC / TT |
|
|
CT + CC / TT |
|
|
CC / CT + TT |
|
|
|
|
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
或(95%置信区间) |
Ph |
P或 |
总计 |
7 |
1.032 (0.922, 1.156) |
0.266 |
0.583 |
0.967 (0.806, 1.161) |
0.688 |
0.720 |
1.017 (0.914, 1.132) |
0.597 |
0.752 |
0.939 (0.789, 1.117) |
0.351 |
0.477 |
种族 |
高加索人 |
6 |
1.061 (0.940, 1.197) |
0.304 |
0.339 |
0.984 (0.813, 1.190) |
0.615 |
0.865 |
1.045 (0.932, 1.171) |
0.716 |
0.454 |
0.947 (0.790, 1.134) |
0.254 |
0.553 |
亚洲 |
1 |
0.846 (0.610, 1.173) |
- |
0.316 |
0.806 (0.430, 1.511) |
- |
0.501 |
0.839 (0.616, 1.143) |
- |
0.266 |
0.853 (0.460, 1.585) |
- |
0.616 |
控制源 |
PB |
4 |
1.045 (0.907, 1.203) |
0.561 |
0.547 |
1.019 (0.818, 1.269) |
0.306 |
0.867 |
1.040 (0.910, 1.189) |
0.737 |
0.564 |
0.990 (0.804, 1.218) |
0.192 |
0.922 |
乙肝 |
3. |
0.895 (0.724, 1.106) |
0.878 |
0.305 |
0.922 (0.632, 1.346) |
0.840 |
0.674 |
0.898 (0.734, 1.098) |
0.846 |
0.293 |
0.964 (0.669, 1.390) |
0.837 |
0.845 |
样本大小 |
≥500 |
5 |
0.997 (0.883, 1.126) |
0.588 |
0.963 |
0.999 (0.821, 1.214) |
0.578 |
0.990 |
0.996 (0.888, 1.118) |
0.671 |
0.948 |
0.987 (0.820, 1.189) |
0.457 |
0.892 |
< 500 |
2 |
1.289 (0.947, 1.756) |
0.112 |
0.107 |
0.779 (0.466, 1.302) |
0.624 |
0.341 |
1.167 (0.870, 1.566) |
0.259 |
0.302 |
0.675 (0.413, 1.101) |
0.315 |
0.115 |
系统回顾-765G>C与CRC易感性之间的关系
总体而言,多态性-765G>C在四种遗传模式下均不影响CRC易感性。meta分析中涉及的研究由于种族组成的多样性、对照来源的差异、遗传背景的差异以及样本量的变化而存在异质性,这些都可能改变最终的结果。因此,进行亚组分析,以考虑混杂因素的潜在影响。对于-765G>C多态性,仅在杂合子和显性模式下的亚洲后裔亚组中存在显著的CRC风险(CG vs. GG, OR=1.477, 95% CI= 1.148-1.898,Ph= 0.242,P或= 0.002;CG+CC vs. GG, OR=1.451, 95% CI=1.146-1.747,Ph= 0.387,P或=0.001)(图5 A-B,表S3)。
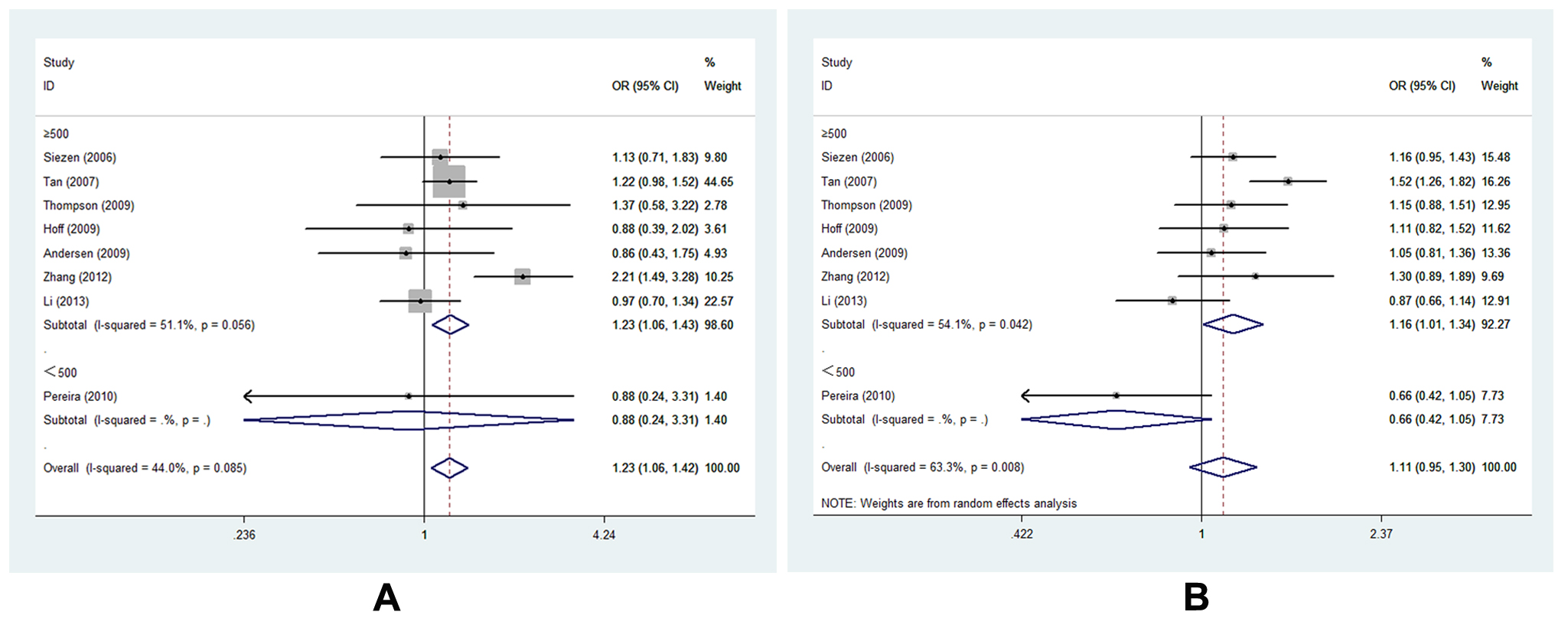
图4。在不同遗传模型样本量亚组分析中,COX-2 -1195G>基因多态性与CRC易感性相关:A, -1195G>A基因的森林图(AG vs GG, size≥500);B, -1195G>A森林地块(AA vs. AG+GG,面积≥500)

图5。不同遗传模式种族亚组分析中COX-2 -765G>C多态性与CRC易感性相关的森林图:A, -765G>C的森林图(CG vs. GG);B, -765G>C的森林分布图(CG+CC vs. GG)。
8473T>C与CRC风险相关性的meta分析
8473T>C多态性与CRC风险之间的相关性没有在整体上或任何子组中被确定(表S3)。
系统综述环境因素和COX-2多态性对CRC易感性的相互影响
在16项符合条件的研究中,只有5篇文章报道了环境因素和COX-2多态性对CRC易感性的潜在交互影响[17,21,22,24,27]。更重要的是,结果是有争议的(表S4)。以NASID为例,一项研究报告,COX-2 -1195AA基因型[27]的患者使用非甾体抗炎药可降低CRC风险。另一篇论文证明了这种关系的负面结果[21]。吸烟的情况也类似。吸烟相关风险在-765GG基因型人群中更强,显示吸烟者发生结直肠癌的风险是不吸烟者[17]的2.682倍。相比之下,Daraei的[24]研究结果为阴性。在此,我们基于有限的可用数据进行元分析,以估计精确的交互效应。结果显示-765GG基因型吸烟者比非吸烟者更容易发生结直肠癌(OR=1.761, 95% CI=1.144-2.711,Ph= 0.286,P或=0.010)(图6,表S4)。

图6。-765GG基因型结直肠癌吸烟相关风险的森林图。通过生成lnor、lnll和lnull参数,使用效果/CI组合进行meta分析。
表4。COX-2基因多态性与环境因素相互影响CRC风险的OR和P值总结
口服补液盐,优势比;cox - 2, cyclooxygenase-2;CRC,结直肠癌;CI,置信区间;异质性检验P值<0.05时,采用随机效应模型;否则,采用固定模型。
-1195 g > |
非甾体抗炎药的使用 |
AA(95%置信区间) |
GA + GG(95%置信区间) |
P价值 |
结合P价值 |
安徒生[21] |
没有 |
裁判 |
0.943 (0.689, 1.292) |
- |
0.715 |
|
|
是的 |
1.020 (0.728, 1.428) |
1.004 (0.659, 1.531) |
0.908 |
0.984 |
|
李[27] |
没有 |
裁判 |
1.08 (0.78, 1.49) |
- |
> 0.05 |
|
|
是的 |
0.19 (0.04 - -0.87) |
0.58 (0.22, 1.50) |
< 0.05 |
> 0.05 |
0.734 |
-1195 g > |
吸烟情况 |
AA(95%置信区间) |
GA + GG(95%置信区间) |
P价值 |
结合P价值 |
佩雷拉[22] |
没有 |
裁判 |
0.776 (0.405, 1.488) |
- |
0.445 |
|
|
是的 |
0.659 (0.354, 1.226) |
6.085 (2.322, 15.95) |
0.188 |
0.000 |
|
李[27] |
没有 |
裁判 |
1.50 (1.02, 2.22) |
- |
> 0.05 |
|
|
是的 |
1.26 (0.77 - -2.06) |
1.08 (0.72, 1.62) |
> 0.05 |
> 0.05 |
|
0.077 |
-765 g > C |
非甾体抗炎药的使用 |
GG(95%置信区间) |
GC + CC(95%置信区间) |
P价值 |
结合P价值 |
安徒生[21] |
没有 |
裁判 |
1.094 (0.775, 1.546) |
- |
0.609 |
|
|
是的 |
1.135 (0.830, 1.553) |
0.876 (0.542, 1.414) |
0.426 |
0.587 |
|
Daraei [24] |
没有 |
裁判 |
1.933 (1.067, 3.501) |
- |
0.030 |
|
0.125 |
|
是的 |
0.994 (0.341, 2.896) |
0.598 (0.232, 1.538) |
0.991 |
0.286 |
|
-765 g > C |
吸烟情况 |
GG(95%置信区间) |
GC + CC(95%置信区间) |
P价值 |
结合P价值 |
邢[17] |
没有 |
裁判 |
0.618 (0.250, 1.526) |
- |
0.297 |
|
|
是的 |
2.682 (1.336, 5.385) |
2.265 (0.894, 5.734) |
0.006 |
0.085 |
|
Daraei [24] |
没有 |
裁判 |
1.548 (0.872, 2.746) |
- |
0.135 |
|
|
是的 |
1.153 (0.288, 4.609) |
1.681 (0.693, 4.080) |
0.840 |
0.251 |
0.010 |
发表偏倚
采用漏斗图、Begg检验和Egger检验对所有难读文章的发表偏倚进行评价。漏斗图的对称形状(图7 A-D)表明所有多态性分析均不存在明显的发表偏倚。此外,Begg 's检验和Egger 's检验的计算结果在统计学上验证了阴性结果(图8)。

图7。发表偏倚检验漏斗图:A, -1195G>A的漏斗图(AG vs. GG);B, -1195G>A漏斗图(AG vs GG,尺寸≥500);C, -1195G>A漏斗图(AA vs. AG+GG,尺寸≥500);D, -765G>C (CG vs. GG,亚洲人)和-765G>C (CG+CC vs. GG,亚洲人)漏斗图;每一个点代表这个更新的元分析中涉及的一个单独的研究;E, Begg漏斗图和Egger发表偏倚图P值的统计分析。
讨论
结直肠癌(CRC)是世界上第三大常见恶性肿瘤,是由遗传变异和环境因素长期不平衡变化引起的。炎症被认为是CRC[3]的一个关键危险因素。COX-2作为一种促炎酶,越来越多的研究探讨了COX-2在结直肠癌发生和发展中的潜在作用,证明了其在结直肠癌发生中的重要作用[5,6,35]。COX-2的功能性遗传变异导致COX-2酶表达和/或活性的改变,可能影响个体发展为CRC[36]的风险。据报道,在COX-2启动子区发现的-1195A>G和-765G>C多态性[37,38],以及在AU-rich元素区(3'UTR)发现的8473T>C多态性可能通过对COX-2的调节作用改变CRC发病的遗传易感性[39-41]。然而,结果却是矛盾的。在这种情况下,我们进行了目前的meta分析,包括12项关于-765G>C的研究[12-14,16-24],8项关于-1195G> a的研究[15,16,19-22,25,27],以及7项关于8473T>C的研究[13,15,19,21,25,26],以阐明这三种多态性对CRC易感的潜在影响。基于本meta分析的结果,我们认为-1195G>A多态性与世界范围内的CRC易感性显著相关,而-765G>C仅在亚洲人群中是潜在的CRC危险因素。此外,8473T>C多态性对CRC易感性无影响。
尽管COX-2 -1195G>A和-765G>C多态性对CRC发病的作用机制尚未明确阐明,但多项研究表明-1195G>A的变异A等位基因和-765G>C的变异C等位基因可以增加COX-2的表达和活性[37,38]。有趣的是,我们发现-765G>C多态性只在亚洲后裔中增加CRC易感性,这与之前在高加索或荷兰人群中的研究[13,20]相反。-765C携带者在不同种族中对CRC发育的不同影响有多种原因。首先,不同的民族拥有不同的遗传背景。在本meta分析中,亚裔和白种人在对照组中C等位基因的频率有显著差异,亚裔为0.03,白种人为0.16。其次,吸烟等环境因素可增加特定基因型个体的CRC风险[17,22]。第三,已有研究报道NSAIDs可通过抑制COX-2酶活性和前列腺素合成降低结直肠癌风险[9,10,42]。与亚洲相比,欧洲和美国是非甾体抗炎药的最大消费国。因此,COX-2 -765G>C多态性在亚洲人群中的风险效应可能更明显。8473T>C多态性被认为是调节COX-2表达的潜在候选基因,其位置位于COX-2的3′-非翻译区(UTR),通过改变mRNA的稳定性和降解来调节COX-2表达[43-45]。 However, 8473T>C polymorphism was not associated with CRC occurrence in this meta-analysis, which was in accordance with previously published studies [13,15,19,21,22,25]. In contrast, it was reported that 8473T>C polymorphism tremendously affected the genetic susceptibility for other epithelial malignancies, such as prostate cancer [46], lung cancer [47]. These findings seemed to suggest that the role of 8473T>C polymorphism was different or even opposite in different types of cancer.
基因背景和环境因素之间的相互影响被认为是CRC发展的根本原因[34]。一项研究报告,COX-2 -1195AA基因型[27]患者使用非甾体抗炎药可降低结直肠癌风险。此外,吸烟相关风险在-765GG基因型人群中更强,显示吸烟者发生CRC的风险是不吸烟者[17]的2.682倍。然而,其他研究报告了相反的结果[21,24]。在此,我们用元分析的方法来评价综合效应。基于两项在亚洲人群中相同多态性的已有研究[17,24],我们认为-765GG基因型的吸烟者比非吸烟者更容易发生结直肠癌。然而,nasid与-1195G>A和-765G>C多态性之间没有关联,因为每个多态性只有两篇论文,一篇是亚洲人的,一篇是高加索人的。因此,对结果的解释应谨慎,并应进行更多考虑基因-基因和基因-环境相互作用的不同种族的研究,以使研究结果有效。
在这个系统的审查中,不存在出版偏倚。但是,有几个限制需要提到。首先,纳入研究的样本量从230急剧偏离到2300。小样本量的研究可能没有足够的统计力量来确定特定多态性和CRC风险之间的真实关联。其次,对照和病例的基线,如年龄、性别、非甾体抗炎药使用情况、吸烟状况等,在每项研究中没有统一调整,这可能会产生混杂偏倚。目前的meta分析是基于未调整的or进行的。因此,对meta分析的结果应谨慎解释,并应使用充分的可用数据进行更严格的分析。再次,本meta分析虽然估计了基因-环境相互作用对CRC风险的影响,但由于现有研究有限,难以得出强有力的结论。迫切需要更多的研究来提高统计效力。
总之,-1195G>A多态性在世界范围内促进了CRC易感性,而-765G>C多态性显著影响了亚洲后裔的CRC风险。-765GG基因型吸烟者更容易发生结直肠癌。进一步考虑基因-基因和基因-环境相互作用的大规模研究需要验证这些发现。
致谢
我们感谢所有对论文给予技术支持和有益讨论的人。
参考文献
- Jemal A, Bray F, Center MM, Ferlay J, Ward E,等(2011)全球癌症统计。CA癌症J临床61: 69 - 90。[Crossref]
- Siegel R, Naishadham D, Jemal A(2013)癌症统计,2013。CA癌症J临床63: 11-30。[Crossref]
- 王东,Dubois RN (2010) COX-2在肠道炎症和结直肠癌中的作用。致癌基因29日:781 - 788。[Crossref]
- Smith WL, Langenbach R(2001)为什么有两种环氧合酶同工酶。中国投资107: 1491 - 1495。[Crossref]
- 姚亮,刘峰,洪亮,孙亮,梁珊,等(2011)COX-2在胃癌细胞血管生成中的作用及机制。临床癌症研究进展30: 13。[Crossref]
- 王艳红,吴文明,杨阿克,张卫东,孙娟等(2011)COX-2基因通过VEGF-C途径促进舌癌细胞增殖和侵袭。医学杂志28配套1:S360-366。[Crossref]
- Elzagheid A, Emaetig F, Alkikhia L, Buhmeida A, Syrjänen K等(2013)环氧化酶-2高表达与结直肠癌晚期相关。抗癌物33: 3137 - 3143。[Crossref]
2021年版权燕麦。所有权利reserv- 彭丽,周燕,王燕,Mou H,赵强(2013)结直肠癌COX-2免疫组化表达的预后意义:文献荟汇分析。《公共科学图书馆•综合》8: e58891。[Crossref]
- Dube C, Rostom A, Lewin G, Tsertsvadze A, Barrowman N, et al.(2007)阿司匹林在结直肠癌一级预防中的使用:为美国预防服务工作小组准备的系统综述。安实习生地中海146: 365 - 375。[Crossref]
- Baron JA, Cole BF, Sandler RS, Haile RW, Ahnen D,等(2003)阿司匹林预防结直肠腺瘤的随机试验。N英语J医学348: 891 - 899。[Crossref]
- Brown JR, DuBois RN (2005) COX-2:预防结直肠癌的分子靶点。中华肿瘤防治杂志23日:2840 - 2855。[Crossref]
- Hamajima N, Takezaki T, Matsuo K, Saito T, Inoue M, et al.(2001)日本结直肠癌患者和非结直肠癌患者环氧酶2 (COX2)罕见多态性的基因型频率。亚洲癌症预防2: 57 - 62。[Crossref]
- Cox DG, Pontes C, Guino E, Navarro M, Osorio A,等。(2004)前列腺素合成酶2/环氧合酶2 (PTGS2/COX2)多态性与结直肠癌的风险。Br J癌症91: 339 - 343。[Crossref]
- Siezen CL, Bueno-de-Mesquita HB, Peeters PH, Kram NR, van Doeselaar M,等(2006)花生四烯酸途径、鱼类摄入和结直肠癌风险相关基因的多态性。Int J癌症119: 297 - 303。[Crossref]
- Koh WP,袁建明,van den Berg D, Lee HP, Yu MC(2004)环氧化酶-2基因多态性与饮食中n-6多不饱和脂肪酸在结肠癌风险中的相互作用:新加坡华人健康研究。Br J癌症90: 1760 - 1764。[Crossref]
- 谭伟,吴娟,张旭,郭燕,刘娟,等(2007)结直肠癌环氧合酶-2和血小板12-脂氧合酶功能多态性与发生风险和晚期疾病状态的关系。致癌作用28日:1197 - 1201。[Crossref]
- 邢丽丽,王宗忠,蒋玲,张燕,徐云云,等(2008)环氧合酶2多态性与结直肠癌:-765G>C基因改变吸烟和体重指数相关风险。世界杂志14: 1785 - 1789。[Crossref]
- Iglesias D, Nejda N, Azcoita MM, Schwartz S Jr, González-Aguilera JJ等(2009)COX2 -765G>C和C . 3618a >G基因多态性对散发性结直肠癌风险和生存的影响。癌症导致控制20: 1421 - 1429。[Crossref]
- Thompson CL, Plummer SJ, Merkulova A, Cheng I, Tucker TC,等(2009)环氧化酶-2和尿苷二磷酸葡萄糖醛酸基转移酶1A6基因多态性与结肠癌风险无相关性。世界杂志15: 2240 - 1144。[Crossref]
- Hoff JH, te Morsche RH, Roelofs HM, van der Logt EM, Nagengast FM等(2009)COX-2多态性-765G—>C和-1195A—>G与结直肠癌风险。世界杂志15: 4561 - 4565。[Crossref]
- Andersen V, Ostergaard M, Christensen J, Overvad K, Tjønneland A,等(2009)在丹麦的一项前瞻性病例队列研究中,异种生物转运体多药耐药1 (MDR1)的多态性和与肉类摄入的相互作用与结直肠癌风险的关系。BMC癌症9: 407。[Crossref]
- Pereira C, pimenel - nunes P, Brandão C, Moreira-Dias L, Medeiros R,等(2010)COX-2多态性与结直肠癌风险:化学预防策略。欧洲J胃肠醇肝醇22日:607 - 613。[Crossref]
- 王杰,Joshi AD, Corral R, Siegmund KD, Marchand LL,等(2012)致癌物代谢基因、红肉和家禽摄入量与结直肠癌风险。Int J癌症130: 1898 - 1907。[Crossref]
- Khorshidi F, Haghighi MM, Nazemalhosseini Mojarad E, Azimzadeh P, Damavand B等(2012)伊朗人群中PTGS2 (COX2) -765G>C基因多态性与散发性结直肠癌的风险。亚洲癌症预防15: 3507 - 11所示。[Crossref]
- 张燕,刘春春,彭海萍,张建忠,蔡晓强,等。(2012)COX-2基因启动子区多态性与结直肠癌易感性的关系。世界华人J消化醇20: 1579 - 1584。
- Ross J, Lockett L, Brookes D, Tabor B, Duesing K,等(2013)PTGS2 rs5275多态性与遗传非综合征易感性家庭结直肠癌风险的关系。Eur J Hum Genet21日:1389 - 1395。[Crossref]
- 李松,赵旭,吴铮,李艳,朱磊,等(2013)花生四烯酸代谢相关基因多态性与结直肠癌的风险和预后。Fam癌症12: 755 - 765。[Crossref]
- Cochran WG(1954)来自不同实验的估计的组合。生物识别技术10: 101 - 129。
- Higgins JP, Thompson SG, Deeks JJ, Altman DG(2003)测量元分析中的不一致性。BMJ327: 557 - 560。[Crossref]
- Mantel N, Haenszel W(1959)疾病回顾性研究数据分析的统计方面。美国国立癌症研究所22日:719 - 748。[Crossref]
- DerSimonian R, Laird N(1986)临床试验的meta分析。控制中国新药临床试验7: 177 - 188。[Crossref]
- Begg CB, Mazumdar M(1994)发表偏倚的秩相关检验的运行特征。生物识别技术: 1088 - 1101。[Crossref]
- Egger M, Davey Smith G, Schneider M, Minder C(1997)通过简单的图形测试检测元分析中的偏倚。BMJ315: 629 - 634。[Crossref]
- 王晓燕,王晓燕,王晓燕,等。(2005)大肠癌。《柳叶刀》365: 153 - 165。[Crossref]
- Mariani F, Sena P, Marzona L, Riccio M, Fano R, et al.(2009)环氧合酶-2和缺氧诱导因子-1 α蛋白表达与炎症相关,且自结直肠癌发生早期上调。癌症列托人279: 221 - 9。[Crossref]
- Pereira C, Medeiros RM, Dinis-Ribeiro MJ(2009)胃癌和结直肠癌中的环氧合酶多态性:是否有确凿的结果?欧洲J胃肠醇肝醇21日:76 - 91。[Crossref]
- Papafili A, Hill MR, Brull DJ, mccanulty RJ, Marshall RP,等(2002)环氧合酶-2常见启动子变异抑制基因表达:在急性期炎症反应中作用的证据。动脉粥样硬化血栓血管生物学22日:1631 - 1636。[Crossref]
- 张旭,苗霞,谭伟,宁斌,刘震,等(2005)环氧化酶-2基因功能变异的鉴定及其与食管癌风险的关系。胃肠病学129: 565 - 576。[Crossref]
- Dixon DA, Kaplan CD, McIntyre TM, Zimmerman GA, Prescott SM(2000)环氧合酶-2基因表达的转录后控制。3'-未翻译区域的作用。J临床生物化学275: 11750 - 11757。[Crossref]
- Langsenlehner U, Yazdani-Biuki B, Eder T, Renner W, Wascher TC,等(2006)环氧化酶-2 (PTGS2) 8473T>C多态性与乳腺癌风险相关。中国癌症Res12: 1392 - 1394。[Crossref]
- Campa D, Zienolddiny S, Maggini V, Skaug V, Haugen A,等(2004)环加氧酶2基因常见多态性与非小细胞肺癌风险的关系。致癌作用25日:229 - 235。[Crossref]
- Gupta RA, Dubois RN(2001)抑制环氧合酶-2预防和治疗结直肠癌。Nat牧师癌症1:乳。[Crossref]
- Cok SJ, Morrison AR.(2001)小鼠环氧合酶-2的3 ' -非翻译区包含多种调节元件,可改变信息的稳定性和翻译效率。J临床生物化学276(25): 23179 - 85。[Crossref]
- Moore AE, Young LE, Dixon DA(2012)环氧合酶-2中常见的单核苷酸多态性破坏了microrna介导的调控。致癌基因31日:1592 - 1598。[Crossref]
- Caput D, Beutler B, Hartog K, Thayer R, Brown-Shimer S, et al.(1986)在指定炎症介质的mRNA分子的3 ' -非翻译区识别一个共同的核苷酸序列。美国国家科学研究院83: 1670 - 4。[Crossref]
- Mandal RK, Mittal RD(2011)在印度北部队列中COX-2基因多态性影响前列腺癌易感性。地中海拱形物42: 620 - 626。[Crossref]
- Park JM, Choi JE, Chae MH, Lee WK, Cha SI,等(2006)环氧合酶8473T>C多态性与肺癌风险的关系:一项病例-对照研究。BMC癌症6: 70。[Crossref]







