摘要
虽然上皮性卵巢癌(EOCs)最初用铂基化疗治疗,但EOCs对铂的反应性不同。根据对铂耐药和铂敏感的EOC细胞系的药效对抗肿瘤药物进行分类,对于开发避免铂无效和开发铂敏感性的治疗策略是有价值的。以未表达FANCF的TOV-21G、OV-90和SKOV-3分别作为铂敏感、铂中间和铂耐药细胞系的例子。抗肿瘤药物检查包括丝裂霉素C,阿霉素,依托泊苷,吉西他滨,氯霉素,紫杉醇,曲平和x射线。通过克隆法分析它们对细胞系的有效性。丝裂霉素C和卡铂的细胞毒性图谱相似,丝裂霉素C对TOV-21G表现出比卡铂更大的效力和选择性。阿霉素、乙泊苷和x射线的细胞毒性图谱与卡铂重叠,而过表达Rad51的OV-90对氯霉素的耐药性高于SKOV-3。紫杉醇和三氮平的疗效与铂敏感性或耐药无关。与这些细胞毒性谱一致的是,顺铂/丝分裂霉素C、三平和紫杉醇诱导H2AX磷酸化的能力不同,并在HL-60人类白血病细胞中产生独特的DNA/RNA合成抑制模式。紫杉醇和三氮平联合对M109小鼠肺癌产生附加的抗肿瘤作用。 In conclusion, mitomycin C is potentially more effective against Fanconi anemia pathway-deficient EOCs than carboplatin. Doxorubicin and etoposide, because of their overlapping cytotoxic properties with carboplatin, are unlikely to be efficacious against platinum-refractory EOCs. Paclitaxel and triapine are effective regardless of platinum sensitivity status, and promising in combination for both platinum-sensitive and platinum-refractory EOCs.
关键字
卡铂,范可尼贫血途径,丝裂霉素C,紫杉醇,铂反应性,三氮平
简介
上皮性卵巢癌(EOC)虽然发病率低,但由于直到疾病发展到晚期才出现症状,因此被称为“无声杀手”。EOC的标准初始处理包括最大限度的去体积手术,然后以铂为基础的化疗。大多数患者对一线治疗有完全的反应。然而,晚期疾病患者无一例外地发生铂敏感性或铂难降解性复发,定义为分别在诱导治疗停止后6个月以上或6个月内复发[1]。这些临床表现与原发性EOCs对铂反应的异质性一致。
事实上,顺铂已被同样有效、肾毒性小得多的类似物卡铂[2]所取代。这些试剂通过DNA镀镀产生链内(占总加合物的90%)和链间(占总加合物的一小部分)交联[3],从而产生细胞毒性。虽然对顺铂的获得性耐药可能涉及多因素过程[4],但一些观察表明,个体肿瘤中的DNA修复能力在决定对铂化合物的固有敏感性方面起着关键作用。例如,来自范可尼贫血(FA)和干皮色素沉着症(XP)患者的成纤维细胞对顺铂[5]同样敏感。多种DNA修复因子决定细胞对顺铂的反应性[6,7]。
种系BRCA1/2突变的遗传性卵巢癌的特征是铂敏感性和总生存率提高[8]。BRCA1 (FANCS)和BRCA2 (FANCD1)属于一组至少16个基因,其中双等位基因突变使个体倾向于fa[9]。此外,FA基因中有害的种系和/或体细胞突变,如BRIP1 (FANCJ)、PALB2 (FANCN)和RAD51C (FANCO)已被鉴定为插入性和非浆液性EOCs[10]。FA通路对修复DNA链间交联(ICLs)至关重要。FA基因产物形成多亚基单泛素连接酶和协同na修复过程,包括xpf - ercc1和Mus81-Eme1的溶核切口、翻译合成和同源重组[11]。根据癌症基因组图谱(TCGA)研究网络的报告,大约一半的高浆液性EOCs[12]存在同源重组缺陷。
XP具有对紫外线照射极度敏感的特征,其原因是核苷酸切除修复(NER)[13]缺乏。真核NER分为全局基因组NER (GG-NER)和转录偶联NER (TC-NER)[14]。对顺铂的过敏仅限于tc - ner缺陷细胞[15]。此外,据报道,TCGA数据集中8%的高级别浆液性EOCs表现出NER改变[16]。
在brca1或BRIP种系杂合子突变引起的遗传性EOCs中,肿瘤组织中野生型等位基因丢失,突变等位基因保留,而正常组织对突变保持杂合子状态[17,18]。因此,这些肿瘤被认为是DNA修复缺陷的,而个体的正常组织是DNA修复良好的。在家族性BRCA2突变的细胞系模型中,DLD1- brca2dex11 /Dex11 (BRCA2 null)对卡铂极度敏感,而DLD1- brca2dex11 /WT (BRCA2单倍缺陷)与DLD1 (BRCA2二倍体)[19]一样耐药。肿瘤和正常组织中DNA修复能力的差异为icl致药发挥肿瘤选择性提供了广阔的治疗窗口。
在过去的20年里,EOCs患者在诊断后1年、3年和5年生存率几乎没有改善[20]。尽管种系BRCA1/2突变与更好的短期生存有关,但这种优势随着时间的推移逐渐消失,最终在BRCA1携带者[21]中逆转。由于标准的治疗方法还没有超越基于铂的联合化疗,因此需要改进铂敏感和铂难治EOCs的治疗策略。为此,我们根据其对铂敏感、铂中间和铂耐药的EOC细胞系的有效性对临床活性抗肿瘤药物进行了分类。在本报告中,我们发现丝裂霉素C是一种比卡铂更有效和更有选择性的药物,可以对抗范可尼贫血途径缺陷的EOC细胞系,而紫杉醇和曲匹宁是有效的药物,无论铂的敏感性状态。我们相信这些信息对于开发铂的敏感性以及避免铂无效是有价值的。
材料与方法
EOC细胞系和试剂
SKOV-3、OV-90[22]和TOV-21G[22]分别于2012年1月、2014年5月和2014年9月从American Type Culture Collection (ATCC, Manassas, VA)购买。在这些细胞系中有突变和表观遗传异常的报道[23-26]。1980年从Robert C. Gallo博士那里获得的HL-60人髓系白血病细胞于2014年2月被ATCC鉴定。所有细胞系均培养于添加10%胎牛血清、50单位/ml青霉素、50µg/ml链霉素的DMEM/F12培养基中。
卡铂(Tocris Bioscience, Bristol, UK)由于[27]自结合在浓水溶液中稳定,在25 mM的水中溶解并在室温下黑暗中保存。顺铂(Sigma-Aldrich, St. Louis, MO)在DMSO[28]中溶解相对缓慢,在4 mM的0.9% NaCl中溶解,立即别名并在-70°C保存。解冻的同分体只使用了一次。盐酸阿霉素(Sigma-Aldrich)和盐酸吉西他滨(Sigma-Aldrich)的原液在10 mM的水中制备,并在-20°C保存。Triapine (FW: 195)在水介质[29]中基本不溶,本实验室将Triapine (FW: 195)配制成茴硫酸盐(Triapine ITS, FW: 321),以避免在动物研究中使用有机溶剂。丝裂霉素C(来自日本Kyowa Hakko的礼物)、乙泊苷(Sigma-Aldrich)、氯丁醇(Sigma-Aldrich)、1,2-双(甲基磺酰)-(2-氯乙基)联氨(90CE)[30]和紫杉醇(Alfa Aesar, Ward Hill, MA)的库存溶液在2.5至50 mM的无水DMSO中制成,并在-20°C保存。用DMSO进一步稀释原液,加入到细胞培养中,最终DMSO浓度小于0.05%。x射线使用X-RAD 320(精密x射线,北布兰福德,CT)进行。
克隆生存试验和生长抑制试验
预先确定的细胞数量,在6孔板中经过各种处理后每孔产生20到150个菌落,允许其粘附一夜,暴露于细胞毒性制剂,并培养13天进行菌落繁殖。对于鸟嘌呤O6-氯乙基化剂90CE [30],O6-苄基鸟嘌呤,一种抑制剂O6-甲基鸟嘌呤-DNA甲基转移酶(MGMT)修复90CE产生的DNA烷基病变,被用于生成MGMT-废弃条件[30]。菌落染色和处理如前所述[31]。未处理的SKOV-3、OV-90和TOV-21G细胞的集落形成效率分别约为0.3、0.15和0.25。基于细胞计数的生长抑制试验,使用经各种药剂处理3天的HL-60细胞,按照前面描述的[31]进行。
全细胞裂解液的制备和western分析
EOC细胞在1.5 x 106细胞/ 75厘米2用磷酸盐缓冲盐水(PBS)洗一次胰蛋白酶化细胞,并在200 μ l Laemmli’s样品缓冲液中溶解,在100℃下溶解5分钟。裂解液进一步通过QIAshredder (QIAGEN, Valencia, CA)剪切DNA。提取液经6%或15%聚丙烯酰胺凝胶电泳(PAGE)分离。使用G:BoxiChemi XR (Syngene, Frederick, MD)捕获化学发光图像。购买了以下蛋白质的抗体:p53 (sc-6243, Santa Cruz Biotechnology, Santa Cruz, CA);BRCA2 (MAB2476, R&D Systems, Minneapolis, MN);Rad51 (MS-988,克隆3C10, Thermo Scientific, Fremont, CA);RPA70 (MS-692, Thermo Scientific), PCNA (MS-106,克隆PC10, Thermo Scientific);gH2AX (sc-101696, Santa Cruz);脱氧胞苷激酶(MABC188,克隆9D4, EMD Millipore, Temecula, CA);O6-甲基鸟嘌呤dna甲基转移酶(MAB16200,克隆MT 3.1, EMD Millipore)。
HL-60人白血病细胞中DNA/RNA合成和gh2ax形成的测量
通过加入[甲基-]来测定DNA和RNA的合成3.H]胸腺嘧啶和[5-]3.H]尿苷变成酸不溶的部分,如前所述[31]。通过gh2ax分析,从3 × 10中提取组蛋白6细胞完整,H 0.2 M2所以4并受到西方的分析[31]。
Madison 109小鼠肺癌的体内治疗
动物实验由机构动物护理和使用委员会审查和批准。M109细胞皮下接种1 × 106将0.1 ml PBS中的细胞注入雌性CD2F1小鼠的腹部(查尔斯河实验室,威尔明顿,MA)。在植入后第1天开始治疗。根据卡尺测量的长度和宽度估计肿瘤体积,公式为:V = (l x w2) / 2。
统计和数学分析
克隆和生长抑制试验至少重复三次。LC标准偏差90和集成电路50通常低于平均值的20%信用证90从相交0.1个细胞存活的2个数据点插值值,假设两点之间的部分是线性的。集成电路50使用KaleidaGraph软件(Synergy software, Reading, PA)从逻辑3参数回归分析中得到值。
结果
EOC细胞系间铂敏感性的多样性
您et al。[26]在其对PARP抑制剂rucaparib的敏感性影响因子的研究中报道了39个EOC细胞系在卡铂敏感性方面的差异。的集成电路50(浓度抑制50%)最敏感的细胞系TOV-21G (0.26 μ M)和九个最耐药的细胞系(> 10 μ M)之间的差异超过38倍,其余细胞系对卡铂表现出梯度敏感性(图1A)。伊藤et al。[25]已经发现证据表明,TOV-21G细胞对顺铂的超敏是由于Fanconi贫血(FA)通路的中断,由表观遗传沉默的FANCF基因通过CpG岛甲基化。我们选用了tov - 21g (IC50, 0.26 μ m), ov-90 (ic50, 1.3 μ M)和SKOV-3 (IC50, >10µM)分别作为铂敏感、铂中间和铂耐药卵巢癌细胞系的样本。图1B总结了这些细胞系的细胞/菌落形态和已知特征。
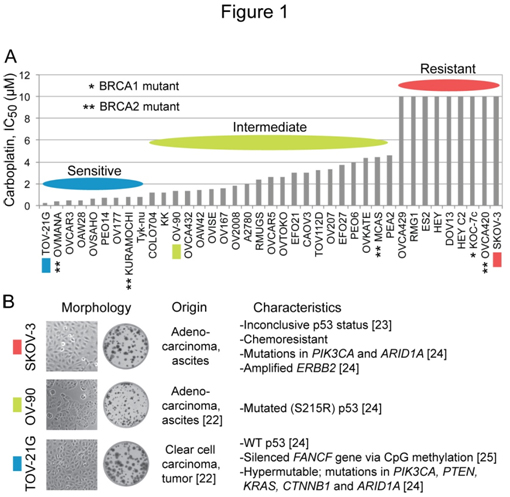
数字1.EOC细胞系卡铂敏感性的多样性,以及研究中使用的SKOV-3、OV-90和TOV-21G细胞的特征。面板A,柱状图是使用报告的IC生成的5039个EOC细胞株[26]中卡铂的值.B组,关于每个细胞系的起源、表型和突变的信息来自ATCC目录和早期的报道[22-25]。
各种抗肿瘤药物对skov -3、OV-90和TOV-21G细胞的作用
三种对铂敏感性不同的EOC细胞系被用于检测卡铂和其他临床活性抗癌药物之间的交叉耐药。图2A显示了每种药剂的浓度依赖性生存曲线。信用证的价值90(浓度给予90%的致死率)以及括号内的折叠抗性值(OV-90和SKOV-3相对于TOV-21G的抗性程度)总结在图2B中。
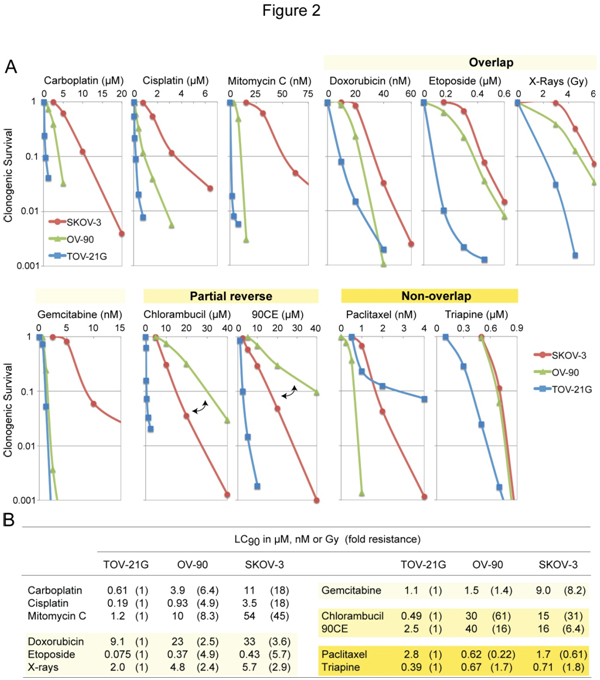
数字2.各种抗肿瘤药物对SKOV-3、OV-90和TOV-21G细胞的细胞毒性研究A组,细胞毒性用克隆法测定。对于鸟嘌呤O6-氯乙基化剂90CE,细胞用10µMO6-苄基鸟嘌呤作用2 h可使其活性减弱O6-甲基鸟嘌呤dna甲基转移酶(MGMT)。O6-苄基鸟嘌呤存在于整个处理过程和群体繁殖过程中。B页,用于澄清LC的值90(浓度给予90%的致死率)和折叠抗性(OV-90和SKOV-3相对于TOV-21G的抗性程度)均列于表中。
尽管碳铂与DNA的反应比顺铂慢得多,但这两种类似物已被证明能产生几乎相同的DNA加合物阵列[32],并产生相似的抗性图谱[33]。与这些发现一致,本研究证实卡铂和顺铂产生高度相似的耐药谱(图2A)。在检测的抗肿瘤药物中,丝裂霉素C产生的敏感性曲线与卡铂产生的最相似(图2A)。此外,丝裂霉素C表现出更大程度的效力(反映在nM的LC90与卡铂的µM,图2B)和选择性(反映在更大的折叠抗性值,图2B)。
相对于卡铂/丝裂霉素C,抗肿瘤药物以重叠、部分倒置和不重叠的方式共同产生敏感性剖面(图2A)。因此,阿霉素、依托泊苷、x射线和吉西他滨产生了重叠的图谱,其排列顺序为TOV-21G O6-氯乙基化剂90CE[30]产生了部分相反的结果,铂中间体OV-90比铂中间体SKOV-3对这些试剂的抗性更强。紫杉醇的排列顺序为OV-90
未处理和药物处理的几种DNA修复/复制蛋白水平
将SKOV-3、OV-90和TOV-21G细胞不处理或用10µM卡铂或1µM三氮平处理24 h。然后,制备全细胞提取物并进行western分析(图3)。三氮平处理后BRCA2和rpa70稳定(不太明显)。三种细胞系之间的显著差异是rad51重组asein OV-90细胞的过表达(图3)。经过三氮平处理后,H2AX (gH2AX)的磷酸化在TOV-21G中显著发生。PCNA水平几乎恒定,无论细胞系和治疗,并作为负荷控制。
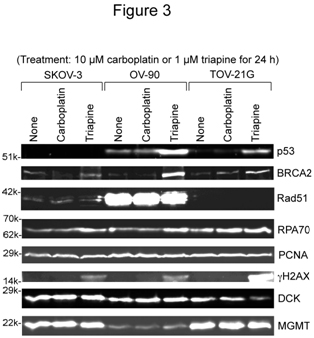
数字3.卡铂或三氮平治疗后相关DNA损伤反应蛋白的西方分析。将未处理的SKOV-3、OV-90和TOV-21G细胞,或用10µM卡铂或1µM triapine处理24 h制备全细胞提取物。BRCA2、p53和RPA70通过6%的PAGE分解。Rad51、PCNA、gH2AX、DCK和MGMT的PAGE分辨率为15%。RPA70,复制蛋白a70 kDa dna结合亚基;增殖细胞核抗原PCNA;脱氧胞苷激酶;管理,O6-methylguanine-DNA甲基转移酶。
版权所有OAT。版权所有在p53状态方面,tov - 21g表现出野生型特征,即在未处理的细胞中未检测到,在三平处理的细胞中积累。在OV-90细胞[24]中报道的S215R突变以及triapine治疗后的显著积累(图3)表明OV-90细胞同时包含野生型和突变的tp53基因。在未处理和三氮平处理的条件下,SKOV-3细胞中p53的缺失表明该细胞系缺乏功能性的altp53基因。
SKOV-3细胞对吉西他滨的耐药性是OV-90和TOV-21G细胞的6- 8倍(图2B)。SKOV-3中较高水平的脱氧胞苷激酶(DCK,负责吉西他滨的初始磷酸化)(图3)排除了作为耐药机制的激活步骤的缺失。
卡铂/丝裂霉素C、三平和紫杉醇对DNA/RNA合成和H2AX磷酸化的抑制作用
DNA链间交联剂剪刀铂和丝裂霉素Cis的细胞毒活性归因于DNA复制和/或转录[4]的干扰,而三平和紫杉醇的细胞毒活性与铂的敏感性/耐药性无关(图2A和2B)。为了深入了解这些独特的细胞毒性特性,我们用HL-60人类白血病细胞测量了这些药物在短期(2小时)和长期(18小时)治疗后抑制DNA和RNA合成的能力。对于每种药剂,选择的最低浓度处理接近其各自的IC50值,由生长抑制试验确定(图4A)。
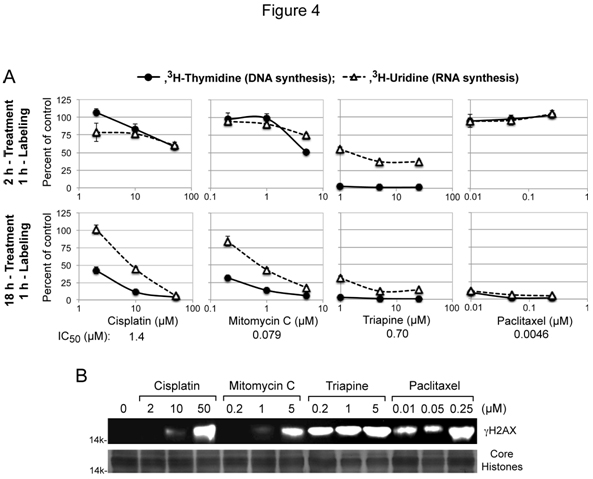
数字4.顺铂、丝裂霉素C、三平或紫杉醇对DNA/RNA合成和gH2AX诱导的抑制作用。面板A,用于短期治疗的HL-60细胞(2 x 106细胞/ml)用各种药剂处理2 h,用3.H-thymidine或3.对于长期治疗,HL-60细胞(4 x 105细胞/ml)用各种药剂处理18 h,浓缩为2 × 106细胞/ml,用3.H-thymidine或3.B组,HL-60细胞(4 x 105细胞/ml)用各种试剂处理18小时。用抗gh2ax抗体对从完整细胞中提取的组蛋白进行15% PAGE和western分析。作为负载对照,使用GelCode Blue安全蛋白染色剂(Thermo Scientific)对核心组蛋白进行重复凝胶染色。
顺铂和丝裂霉素C产生几乎相同的抑制模式,只有在处理18小时后才优先抑制DNA合成(图4A)。相反,在短期(2小时)治疗中,三氮平导致DNA合成立即完全停止(图4A)。紫杉醇对核酸合成的影响不同于三氮平和顺铂/丝分裂霉素c。因此,紫杉醇处理2 h后核酸合成不间断,处理18 h后DNA和RNA合成完全停止(图4A)。
DNA双支架断裂在染色质微环境[34]中诱导组蛋白H2AX(gH2AX)的快速磷酸化。在HL-60细胞中检测顺铂/丝裂霉素C、三平和紫杉醇诱导h2ax形成的能力,这些药物处理18小时(图4B)。在浓度(0.2µM)比其IC低3.5倍时,Triapine可明显形成gH2AX50价值。相比之下,顺铂/丝裂霉素C在浓度为其IC的40 ~ 60倍时诱导明显的H2AX磷酸化50值。紫杉醇,不直接靶向dna,也导致显著的gh2ax积累。这些结果共同表明,H2AX的磷酸化不足以成为细胞毒性的生物标志物。
抗肿瘤药物按其对铂敏感和铂耐药细胞系的有效性和潜在的作用机制分类
根据生存曲线,和LC90如图2A和2B所示,根据抗癌药物对铂敏感的TOV-21G、铂中间的ov -90和铂耐药的SKOV-3的有效性对其进行了分类(图5A)。丝裂霉素C对fancf缺陷的TOV-21G的疗效显著,比卡铂更有效力和选择性,而紫杉醇和三氮平对铂敏感和铂耐药的EOC细胞均有显著疗效。图5B总结了卡铂/丝裂霉素C、三氮平和紫杉醇的主要作用位点和细胞毒机制,形成了它们独特的细胞毒特性的基础。这个问题将在讨论部分进一步讨论。
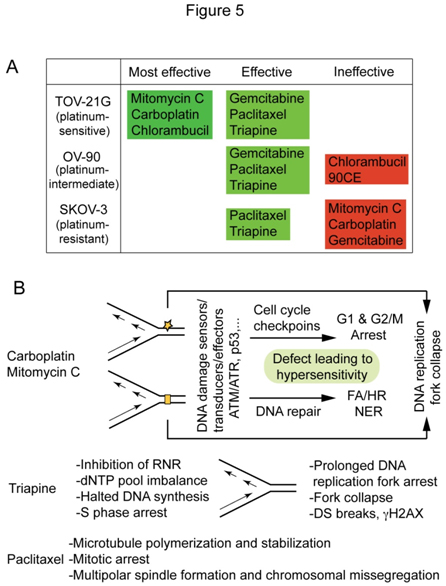
数字5.根据对TOV-21G、OV-90和SKOV-3的药效(A)和卡铂/丝裂霉素C、三平和紫杉醇的细胞毒性机制(B)进行的抗肿瘤药物分类。
紫杉醇单独或联合三氮平对Madison 109小鼠肺癌的抗肿瘤活性
triapine单独或联合顺铂、阿霉素或依托泊苷的有效性已在EOC细胞系A2780异种移植瘤模型[35]中得到证实。在本研究中,我们在M109肺癌(一种对顺铂[36]反应弱的小鼠同基因肿瘤模型)中检测了triapine单独或联合紫杉醇的抗肿瘤活性。图6显示triapine ITS (triapine的水溶性异硫氰酸盐形式)在延缓肿瘤生长方面与紫杉醇一样有效。此外,紫杉醇与三氮平联合使用可增加肿瘤生长的延迟。
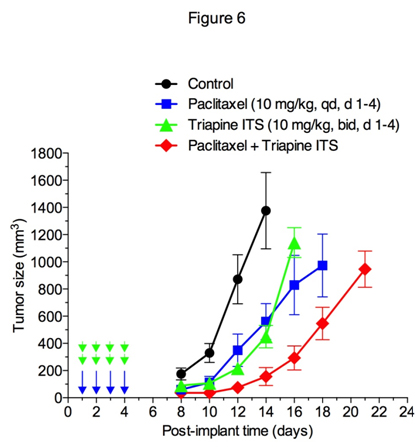
数字6.紫杉醇和三氮平单独或联合使用对Madison 109小鼠肺癌的抗肿瘤活性。荷瘤小鼠(5只/组)腹腔注射紫杉醇10 mg/kg,每日1次,连续4天,异硫酸三氮平盐(triapine ITS) 10 mg/kg(相当于6 mg/kg的三氮平),每日2次,间隔8小时,连续4天,或两者联合使用。用胰岛素注射器按10 μ l/10 g重量注射DMSO溶解的紫杉醇10 mg/ml。以1mg /ml (ph4.5)溶于水中的Triapine ITS用2mol /ml的NaHCO中和3.注射前立即注射。
讨论
尽管含铂方案一直是ocs初始治疗的主要方案,但癌症复发要么是铂难治,要么是铂敏感。因此,有必要(a)理解固有铂耐药性和敏感性的机制,(b)开发方法在化疗前区分铂耐火EOCs和铂敏感EOCs, (c)分别对铂耐火EOCs和铂敏感EOCs应用适当的治疗方法,以避免铂无效和开发铂敏感性。PubMed对顺铂耐药性、顺铂敏感性和顺铂作用机制的搜索结果分别为8400、5500和46000篇文章。尽管有大量的研究,关于DNA加合物(DNA链内或链间交联,或两者)对细胞毒性至关重要的性质,涉及关键DNA加合物去除的DNA损伤修复系统的类型,以及抑制DNA/RNA合成导致最终细胞死亡的一系列事件,仍有很多有待澄清。
Ihnen报道的39个来自浆液性和非浆液性EOCs的EOC细胞系中卡铂敏感性的显著多样性(图1A)等.[26]与NCI-60中适度变化的铂敏感性形成对比,NCI-60是一组来自不同人类癌症的60个细胞系,包括6种卵巢癌(NCI DPT数据)。在39个EOC细胞系中(图1A), 18个细胞系(46%)对卡铂明显敏感(IC50小于2 μ M)。大量铂超敏EOC细胞系的存在与人类原发性EOCs中涉及HR、FA和NER的DNA修复途径的高突变率兼容[10,12,16]。因此,确定这些eeoc细胞系铂超敏的分子机制及其对一系列抗肿瘤药物的敏感性具有重要意义。
在本研究中,我们分别以SKOV-3、OV-90和TOV-21G作为铂耐、铂中间和铂敏感的EOC细胞株为例。根据原发性高级别浆液性卵巢癌(HGSOC)和永久性卵巢癌细胞系的基因组图谱,OV-90和SKOV-3分别可能和不可能来自HGSOC[24]。从透明细胞癌[22]中建立的TOV-21G,已知由于基因沉默导致FANCF表达缺失,并且对顺铂[25]过敏。
细胞对DNA链间交联剂的超敏性是FA[11]的典型特征之一。fancf缺陷的TOV-21G一贯表现出对丝裂霉素C (LC90, 1.2 nM),氯霉素(LC90, 0.49 μ M)和卡铂(LC90, 0.61 μ m)。TOV-21G对丝裂霉素C、氯霉素和卡铂的敏感性分别是耐铂的SKOV-3的45倍、31倍和18倍。更大的效力(LC90丝裂霉素C表现出的选择性(折叠敏感性)表明该制剂对FA通路缺陷EOCs的潜在效用。
值得注意的是,铂中间体OV-90比铂中间体SKOV-3对氯霉素和90CE的抗性更强(部分相反,图2A)。虽然顺铂、丝裂霉素C、氯霉素和90ce4均可引起edna链间交联(ICL),但单个制剂产生的ICL加合物的化学结构非常不同[3,30,37-39]。然而,一方面由顺铂/丝裂霉素C产生的icl,另一方面由氯霉素/90CE产生的icl的修复机制似乎是有区别的。西方对几个关键DNA损伤修复蛋白的分析(图3)清楚地表明OV-90细胞过表达Rad51。由于Rad51与Rad51同源物(Rad51B, Rad51C, Rad51D, XRCC2, XRCC3和减数分裂特异性DMC1)[40]具有相同的氨基酸序列相似性和结构域,因此Rad51同源物在各种icls修复中的作用有待进一步研究。
值得注意的是,吉西他滨对铂敏感的TOV-21G和铂中间体OV-90具有同样的细胞毒性。因此,与阿霉素、依托泊苷和x射线的细胞毒性图谱与卡铂重叠(图2A)不同,吉西他滨的细胞毒性特性似乎是独特的。为了支持这一观点,琼斯等.[41]已经证明功能性BRCA2和Rad51的存在促进(而不是降低)吉西他滨的细胞毒性活性。这些观察结果共同表明吉西他滨对一种未确定的EOCs亚群有效。
本报告的一项重要发现是紫杉醇和三氮平对EOC细胞系的疗效,而不管铂敏感状态,这表明这些药物在治疗铂敏感和铂难治EOCs方面都有用。长期以来,人们已经认识到铂和taxene对steoc的临床疗效及其对steoc细胞系的细胞毒活性呈反比关系,并对其进行了广泛的研究。鉴于这两种药物作用方式的明显差异(图5B),可以想象紫杉醇的细胞毒性活性与铂耐药性或敏感性无关。另一方面,三平和卡铂都是已知的引起DNA复制应激的药物。作为核糖核酸脱氢酶的抑制剂,三氮平的效力是羟基脲[31]的200到300倍,羟基脲[31]是一种历史上用于在培养细胞[43]中产生na复制应激的药剂。图5B显示了由核糖核酸还原酶抑制引起的dNTP池失衡最终传递到DNA双链断裂的顺序事件。卡铂和丝裂霉素c与三氮平的区别在于,这些药物通过直接修饰DNA结构,诱导DNA损伤反应途径,保护细胞免受有害遗传毒性(图5B)。因此,DNA损伤反应途径的缺陷导致细胞对这些制剂的超敏性,而由三氮平引起的大量DNA双支架断裂是次要于长时间DNA复制应激的最终结果(图5B)。
Triapine是在已故Sartorelli博士的实验室[29]中设计和合成的,目前正在nih资助的I/II期临床试验中作为抗肿瘤药物进行评估。选择Triapine进行临床开发是因为其在临床前模型[35]中单独或与顺铂、阿霉素或依托泊苷联合使用具有广谱抗肿瘤活性。使用培养细胞进行的研究表明,由于核糖核酸脱氢酶[31]的抑制导致dNTP池不平衡,浓度为0.4至0.8 μ m的三平会立即停止DNA复制。尽管对DNA合成有瞬时作用,但三氮平对培养细胞的有效杀伤需要较长时间的药物暴露(16小时),这可能是因为三氮平移除后DNA合成的抑制是可逆的,DNA双链断裂发生在长时间的DNA复制应激[31]之后。在向L1210荷瘤小鼠单次注射三氮平后,正常组织(如骨髓和十二指肠)中的DNA合成比肿瘤细胞[35]中恢复得更快。与这些观察结果一致的是,在临床前模型中,triapinea的抗肿瘤活性是时间表依赖性的。因此,一天两次的方案(6mg /kg一天两次,间隔8小时,持续5天,总剂量为60mg /kg)大大优于几乎无效的一天一次方案(24mg /kg一天一次,持续5天,总剂量为120mg /kg),总剂量为[35]的两倍。
大多数triapine2小时的临床试验。每日输注三氮平,剂量高达120 mg/m2I期试验确定的方案为可耐受剂量,这可能不是最佳治疗[31]。应在临床试验中探索可使三氮平血浆浓度保持在较低水平并使DNA复制应激维持较长时间的分剂量方案。此外,低但持续的三氮平浓度是无效的可能引起宿主毒性,如高铁血红蛋白血症和缺氧。
在本研究中,我们根据对铂敏感型TOV-21G、铂中间型OV-90和铂耐型SKOV-3的药效对抗肿瘤药物进行了分类。总的来说,铂的敏感性/耐药性是多因素的。因此,个体化用药的直接方法应该包括体内药敏分析,其中可以测试单个EOC样本对一系列抗肿瘤药物的敏感性/耐药性。有效分析的先决条件包括:a)通过联合阳性(EpCAM珠)和阴性(抗成纤维细胞珠)选择来富集EOC细胞;b)提供生长刺激环境以增强对细胞复制靶向抗肿瘤药物的敏感性;c)使用适当的细胞标记物来区分肿瘤细胞与正常细胞;d)使用足够的细胞毒性标记物来检测细胞毒性。建立这样的分析方法对于成功的个性化医疗至关重要。
作者
k.i设计并发展了这项工作的概念,进行了实验,分析了数据并起草了论文。Y-L.Z。, Z-P.L。和r.z进行了实验并分析了数据。p.g.p., k.s., r.p.b., T.J.R.和A.C.S.对这项工作的概念和设计做出了贡献,分析了数据,并完善了报告。esr对这项工作的概念和设计做出了贡献,对数据进行了分析,完善了报告,获得了资金并给予了最终批准。
确认
我们感谢Bret Halpern博士的有益讨论。
资金
这项工作得到了尤尼斯·肯尼迪·施赖弗国家儿童健康和人类发展研究所K12HD047018补助金(E.S.R.)的部分支持,以及E.S.R.从发现到治疗的补助金
参考文献
- Banerjee S, Bookman M, Gore M(2011)卵巢癌的全身治疗,目前的治疗,最近的进展,和未满足的需求。卵巢癌的新治疗靶点。施普林格,伦敦:1-33。
- Kelland LR(2005)卵巢癌新兴药物。新兴药物专家意见10: 413 - 424。(Crossref)
- Jamieson ER, Lippard SJ(1999)顺铂- dna加合物的结构、识别和加工。化学牧师99: 2467 - 2498。(Crossref)
- Jung Y, Lippard SJ(2007)铂诱导DNA损伤的直接细胞反应。化学牧师107: 1387 - 1407。(Crossref)
- Plooy AC, Van Dijk M, Berends F, Lohman PH(1985)用顺式二胺二氯铂处理的对照和突变人类细胞中诱导的细胞毒性和计划外DNA合成中DNA链间交联的形成和修复(II)。癌症Res45: 4178 - 4184。(Crossref)
- Zdraveski ZZ, Mello JA, Marinus MG, Essigmann JM(2000)多种重组途径定义了细胞对顺铂的反应。化学生物7: 39-50。(Crossref)
- Nojima K, Hochegger H, Saberi A, Fukushima T, Kikuchi K, et al.(2005)多种修复途径介导脊椎动物细胞对化疗交联剂的耐受。癌症Res65: 11704 - 11711。(Crossref)
- Bolton KL, Chenevix-Trench G, Goh C, Sadetzki S, Ramus SJ等(2012)侵袭性上皮性卵巢癌患者BRCA1和BRCA2突变与生存率的关系。《美国医学会杂志》307:382 - 390。(Crossref)
- Wang AT, Smogorzewska A (2015) SnapShot: Fanconi贫血及其相关蛋白。细胞160: 354 - 354. - e1。(Crossref)
- Pennington KP, Walsh T, Harrell MI, Lee MK, Pennil CC等(2014)同源重组基因的种系和体细胞突变预测卵巢癌、输卵管和腹膜癌的铂反应和生存。临床癌症治疗20: 764 - 775。(Crossref)
- Kim H, D'andrea AD(2012)范可尼贫血/BRCA通路对DNA交联修复的调控。基因开发26日:1393 - 1408。(Crossref)
- 癌症基因组图谱研究网络(2011)卵巢癌综合基因组分析。自然474: 609 - 615。(Crossref)
- Lehmann AR(2003年)DNA修复缺陷疾病、色素性干皮病、Cockayne综合征和毛硫代营养不良。Biochimie85: 1101 - 1111。(Crossref)
- Marteijn JA, Lans H, Vermeulen W, Hoeijmakers JH(2014)了解核苷酸切除修复及其在癌症和衰老中的作用。Nat Rev Mol细胞生物学15: 465 - 481。(Crossref)
- Furuta T, Ueda T, Aune G, Sarasin A, Kraemer KH, et al.(2002)转录偶联核苷酸切除修复是人类细胞顺铂敏感性的决定因素。癌症Res62: 4899 - 4902。(Crossref)
- Ceccaldi R, O'connor KW, Mouw KW, Li AY, Matulonis UA,等(2015)具有铂敏感性和PARP抑制剂耐药的上皮性卵巢癌的独特亚群。癌症Res75: 628 - 634。(Crossref)
- Neuhausen SL, Marshall CJ(1994)来自三个brca1连锁家族肿瘤的杂合性缺失。癌症Res54: 6069 - 6072。(Crossref)
- Rafnar T, Gudbjartsson DF, Sulem P, Jonasdottir A, Sigurdsson A,等(2011)BRIP1基因突变导致卵巢癌的高风险。Nat麝猫43: 1104 - 1107。(Crossref)
- hul T, Rago C, Gallmeier E, Brody JR, Gorospe M,等人(2008)人类变异功能注释的同源方差库:在BRCA2上的应用。癌症Res68: 5023 - 5030。(Crossref)
- Vaughan S, Coward JI, Bast RC, Jr., Berchuck A, Berek JS,等(2011)卵巢癌的再思考:改善预后的建议。Nat Rev癌症11: 719 - 725。
- Candido-Dos-Reis FJ, Song H, Goode EL, Cunningham JM, Fridley BL等(2015)BRCA1或BRCA2种系突变与上皮性卵巢癌诊断女性的10年生存率。临床癌症治疗21日:652 - 657。(Crossref)
- Provencher DM, Lounis H, Champoux L, Tetrault M, Manderson EN,等(2000)四种新型上皮性卵巢癌细胞系的表征。体外细胞发育生物学研究36: 357 - 361。(Crossref)
- 王晓燕,王晓燕,王晓燕,王晓燕(2008)人肿瘤细胞系中p53基因突变状态分析。癌症生物学7: 701 - 710。(Crossref)
- Domcke S, Sinha R, Levine DA, Sander C, Schultz N(2013)通过比较基因组图谱评估细胞系作为肿瘤模型。Nat Commun4: 2126 - 2135。
- Taniguchi T, Tischkowitz M, Ameziane N, Hodgson SV, Mathew CG,等(2003)顺铂敏感卵巢肿瘤中Fanconi贫血- brca通路的中断。Nat地中海9: 568 - 574。(Crossref)
- Ihnen M, Zu Eulenburg C, Kolarova T, Qi JW, Manivong K,等。(2013)聚(adp核糖)聚合酶抑制剂rucaparib治疗散发性卵巢癌的治疗潜力。Mol Cancer Ther12: 1002 - 1015。(Crossref)
- Di Pasqua AJ, Kerwood DJ, Shi Y, Goodisman J, Dabrowiak JC(2011)卡铂和奥沙利铂注射液的稳定性是由于自缔合。道尔顿反式40: 4821 - 4825。(Crossref)
- Sundquist WI, Ahmed KJ, Hollis LS, Lippard SJ(1987)顺式和反式二胺二氯铂(II)在二甲基亚砜中的溶解反应。反式[Pt(NH)]的结构表征和DNA结合3.)2(我2所以)Cl) +。Inorg化学26日:1524 - 1528。
- 刘春春,林涛,Sartorelli AC(1992)吡啶-2-羧醛硫代氨基脲氨基衍生物的合成及其抗肿瘤活性。医学化学35: 3672 - 3677。(Crossref)
- Ishiguro K, Lin ZP, Penketh PG, Shyam K, Zhu R,等(2014)triapine及其末端二甲基化衍生物Dp44mT由于铜(II)络合物活性的丧失或增加而导致细胞杀伤的不同机制。生物化学杂志91: 312 - 322。(Crossref)
- 石黑,朱永利,Shyam K, Penketh PG, Baumann RP,等(2010)鸟嘌呤的数量关系O6-烷基病变由蛇毒素产生,肿瘤耐药由O6-alkylguanine-DNA alkyltransferase。生物化学杂志80: 1317 - 1325。(Crossref)
- Knox RJ, Friedlos F, Lydall DA, Roberts JJ(1986)抗癌铂类药物的细胞毒性机制:顺式二胺二氯铂(II)和顺式二胺-(1,1-环丁二羧基lato)铂(II)仅在它们与DNA相互作用的动力学上不同的证据。癌症Res46: 1972 - 1979。(Crossref)
- Rixe O, Ortuzar W, Alvarez M, Parker R, Reed E, et al.(1996)奥沙利铂、四铂、顺铂和卡铂:耐药细胞系和国家癌症研究所抗癌药物筛选面板细胞系的活性谱。生物化学杂志52: 1855 - 1865。(Crossref)
- Turinetto V, Giachino C(2015)组蛋白变体H2AX的多个层面:一种具有多种生物学功能的DNA双链断裂标记。核酸保留区43: 2489 - 2498。
- Finch RA, Liu M, Grill SP, Rose WC, Loomis R,等(2000)三氮平(3-氨基吡啶-2-羧基硫代氨基脲):一种具有广谱抗肿瘤活性的核糖核酸还原酶活性抑制剂。生物化学杂志59: 983 - 991。(Crossref)
- Rose WC (1981) Madison 109肺癌作为筛选抗肿瘤药物模型的评价。癌症治疗代表65: 299 - 312。(Crossref)
- Tomasz M(1995)丝裂霉素C:小,快速和致命(但非常有选择性)。化学生物2: 575 - 579。(Crossref)
- Gruppi F, Hejazi L, Christov PP, Krishnamachari S, Turesky RJ,等。(2015)小牛胸腺DNA和人乳腺癌细胞系中氮芥甲脒嘧啶加合物形成双(2-氯乙基)乙胺的表征。化学Res毒理学28日:1850 - 1860。(Crossref)
- dean AJ, West SC (2011) DNA链间交联修复与癌症。Nat Rev癌症11: 467 - 480。
- Suwaki N, Klare K, Tarsounas M (2011) RAD51在DNA损伤信号、重组修复和肿瘤发生中的作用。Semin细胞发育生物学22日:898 - 905。(Crossref)
- Jones RM, Kotsantis P, Stewart GS, Groth P, Petermann E (2014) BRCA2和RAD51促进双链断裂形成和细胞死亡对吉西他滨的反应。Mol Cancer Ther13: 2412 - 2421。(Crossref)
- Stordal B, Pavlakis N, Davey R(2007)从实验到临床铂和紫杉烷耐药性的系统综述:一种反向关系。癌症治疗33: 688 - 703。(Crossref)
- Fugger K, Chu WK, Haahr P, Kousholt AN, Beck H, et al. (2013) FBH1与MUS81协同诱导复制应激后DNA双链断裂和细胞死亡。Nat Commun4: 1423 - 1430。(Crossref)






